
Aromorphoses in Biological аnd Social Evolution: Some General Rules for Biological and Social Forms of Macroevolution
Aromorphoses in Biological аnd Social Evolution: Some General Rules for Biological and Social Forms of Macroevolution
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- BIOLOGY INVESTIGATORY PROJECT - Cardiovascular Diseases - Coronary Artery Disease PDFDocument50 pagesBIOLOGY INVESTIGATORY PROJECT - Cardiovascular Diseases - Coronary Artery Disease PDFlskumar100% (1)
- Revised Curriculum DPT-UHS (15!09!15)Document294 pagesRevised Curriculum DPT-UHS (15!09!15)Iqra Iftikhar50% (2)
- Physiological Regulation of Arterial Blood PressureDocument36 pagesPhysiological Regulation of Arterial Blood PressureFrancesNo ratings yet
- 2.2 - All Worksheets - Homework Sheets - Extension SheetsDocument20 pages2.2 - All Worksheets - Homework Sheets - Extension SheetslogineNo ratings yet
- Valvular Heart Diseases: Cardiology, Med Ii - 3Rd YearDocument11 pagesValvular Heart Diseases: Cardiology, Med Ii - 3Rd YearMiguel Cuevas DolotNo ratings yet
- Department of Engineering Science Institute of Biomedical EngineeringDocument35 pagesDepartment of Engineering Science Institute of Biomedical Engineeringgary_vlaicu3850No ratings yet
- Cardiac AssessmentDocument48 pagesCardiac AssessmentRatheesh NathNo ratings yet
- CVS Examination ProformaDocument4 pagesCVS Examination ProformaAnantha KrishnanNo ratings yet
- Cag Views PPT Sunil PDFDocument57 pagesCag Views PPT Sunil PDFNikul J PanchalNo ratings yet
- EECP IabpDocument16 pagesEECP IabpMythily VedhagiriNo ratings yet
- Anatomy & PhysiologyDocument38 pagesAnatomy & PhysiologyParsa EbrahimpourNo ratings yet
- Pathophysiology - Pleural EffusionDocument1 pagePathophysiology - Pleural Effusionfaula rocamora80% (5)
- GEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Document202 pagesGEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Shantanu MandalNo ratings yet
- Section 5. Atherosclerotic Peripheral Vascular Disease and Aortic AneurysmDocument23 pagesSection 5. Atherosclerotic Peripheral Vascular Disease and Aortic AneurysmsidoniavidicanNo ratings yet
- Acute Fluid Management of Large Burns Pathophysiology, Monitoring, and ResuscitationDocument9 pagesAcute Fluid Management of Large Burns Pathophysiology, Monitoring, and ResuscitationEduardo Espin LanzNo ratings yet
- Myocardial InfarctionDocument4 pagesMyocardial Infarctionجعفر محمدNo ratings yet
- Learners Material Module 1 Respiratory ADocument27 pagesLearners Material Module 1 Respiratory AJelly FloresNo ratings yet
- Circulatory System of CockroachDocument18 pagesCirculatory System of Cockroachyayeg rajaNo ratings yet
- New DOCX DocumentDocument2 pagesNew DOCX DocumentPunjabi FootballNo ratings yet
- A Pediatric Echocardiographic Z Score Nomogram For A Developing Country: Indian Pediatric Echocardiography Study - The Z ScoreDocument8 pagesA Pediatric Echocardiographic Z Score Nomogram For A Developing Country: Indian Pediatric Echocardiography Study - The Z ScoreTia mssNo ratings yet
- 3.diagnostic ProceduresDocument17 pages3.diagnostic ProceduresCeleste GalvanNo ratings yet
- Ecg TestDocument67 pagesEcg TestEm KayNo ratings yet
- Rah, Lah, RVH, LVHKDocument12 pagesRah, Lah, RVH, LVHKYuliana AtmayudhaNo ratings yet
- FundamentalsDocument30 pagesFundamentalsRachel PerandoNo ratings yet
- Inotropes in PicuDocument2 pagesInotropes in PicuDijattxNo ratings yet
- University of Northern PhilippinesDocument3 pagesUniversity of Northern Philippines3B NOVIDA, ALEYA G.No ratings yet
- The Circulatory SystemDocument30 pagesThe Circulatory Systemapi-309893409No ratings yet
- The Circulatory System P2Document17 pagesThe Circulatory System P2Kristal CampbellNo ratings yet
- Hot Sitz Bath and Perilite ExposureDocument29 pagesHot Sitz Bath and Perilite ExposureSusan MaglaquiNo ratings yet
- The Cardiovascular System: The Heart: Anatomy and PhysiologyDocument30 pagesThe Cardiovascular System: The Heart: Anatomy and PhysiologyZachary CohenNo ratings yet













































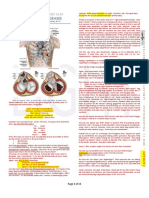






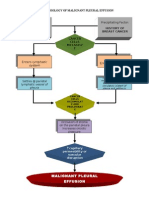










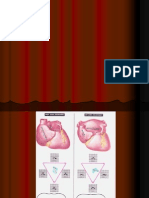







Download as doc, pdf, or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- BIOLOGY INVESTIGATORY PROJECT - Cardiovascular Diseases - Coronary Artery Disease PDFDocument50 pagesBIOLOGY INVESTIGATORY PROJECT - Cardiovascular Diseases - Coronary Artery Disease PDFlskumar100% (1)
- Revised Curriculum DPT-UHS (15!09!15)Document294 pagesRevised Curriculum DPT-UHS (15!09!15)Iqra Iftikhar50% (2)
- Physiological Regulation of Arterial Blood PressureDocument36 pagesPhysiological Regulation of Arterial Blood PressureFrancesNo ratings yet
- 2.2 - All Worksheets - Homework Sheets - Extension SheetsDocument20 pages2.2 - All Worksheets - Homework Sheets - Extension SheetslogineNo ratings yet
- Valvular Heart Diseases: Cardiology, Med Ii - 3Rd YearDocument11 pagesValvular Heart Diseases: Cardiology, Med Ii - 3Rd YearMiguel Cuevas DolotNo ratings yet
- Department of Engineering Science Institute of Biomedical EngineeringDocument35 pagesDepartment of Engineering Science Institute of Biomedical Engineeringgary_vlaicu3850No ratings yet
- Cardiac AssessmentDocument48 pagesCardiac AssessmentRatheesh NathNo ratings yet
- CVS Examination ProformaDocument4 pagesCVS Examination ProformaAnantha KrishnanNo ratings yet
- Cag Views PPT Sunil PDFDocument57 pagesCag Views PPT Sunil PDFNikul J PanchalNo ratings yet
- EECP IabpDocument16 pagesEECP IabpMythily VedhagiriNo ratings yet
- Anatomy & PhysiologyDocument38 pagesAnatomy & PhysiologyParsa EbrahimpourNo ratings yet
- Pathophysiology - Pleural EffusionDocument1 pagePathophysiology - Pleural Effusionfaula rocamora80% (5)
- GEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Document202 pagesGEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Shantanu MandalNo ratings yet
- Section 5. Atherosclerotic Peripheral Vascular Disease and Aortic AneurysmDocument23 pagesSection 5. Atherosclerotic Peripheral Vascular Disease and Aortic AneurysmsidoniavidicanNo ratings yet
- Acute Fluid Management of Large Burns Pathophysiology, Monitoring, and ResuscitationDocument9 pagesAcute Fluid Management of Large Burns Pathophysiology, Monitoring, and ResuscitationEduardo Espin LanzNo ratings yet
- Myocardial InfarctionDocument4 pagesMyocardial Infarctionجعفر محمدNo ratings yet
- Learners Material Module 1 Respiratory ADocument27 pagesLearners Material Module 1 Respiratory AJelly FloresNo ratings yet
- Circulatory System of CockroachDocument18 pagesCirculatory System of Cockroachyayeg rajaNo ratings yet
- New DOCX DocumentDocument2 pagesNew DOCX DocumentPunjabi FootballNo ratings yet
- A Pediatric Echocardiographic Z Score Nomogram For A Developing Country: Indian Pediatric Echocardiography Study - The Z ScoreDocument8 pagesA Pediatric Echocardiographic Z Score Nomogram For A Developing Country: Indian Pediatric Echocardiography Study - The Z ScoreTia mssNo ratings yet
- 3.diagnostic ProceduresDocument17 pages3.diagnostic ProceduresCeleste GalvanNo ratings yet
- Ecg TestDocument67 pagesEcg TestEm KayNo ratings yet
- Rah, Lah, RVH, LVHKDocument12 pagesRah, Lah, RVH, LVHKYuliana AtmayudhaNo ratings yet
- FundamentalsDocument30 pagesFundamentalsRachel PerandoNo ratings yet
- Inotropes in PicuDocument2 pagesInotropes in PicuDijattxNo ratings yet
- University of Northern PhilippinesDocument3 pagesUniversity of Northern Philippines3B NOVIDA, ALEYA G.No ratings yet
- The Circulatory SystemDocument30 pagesThe Circulatory Systemapi-309893409No ratings yet
- The Circulatory System P2Document17 pagesThe Circulatory System P2Kristal CampbellNo ratings yet
- Hot Sitz Bath and Perilite ExposureDocument29 pagesHot Sitz Bath and Perilite ExposureSusan MaglaquiNo ratings yet
- The Cardiovascular System: The Heart: Anatomy and PhysiologyDocument30 pagesThe Cardiovascular System: The Heart: Anatomy and PhysiologyZachary CohenNo ratings yet