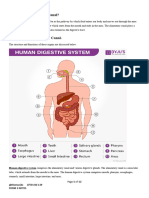
Download as doc, pdf, or txt
You might also like
- Clinical Problem Solving in OrthodonticsDocument294 pagesClinical Problem Solving in OrthodonticsAdina SerbanNo ratings yet
- Dumbbell Exercise Poster 11X17 High ResDocument1 pageDumbbell Exercise Poster 11X17 High ResArturo SierraNo ratings yet
- MODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFDocument27 pagesMODULE 2 Ichthyology - SECTION 5 Shark Anatomy and Dogfish Dissection PDFElishae SamonteNo ratings yet
- Urinary Tract Infection Anatomy and PhysiologyDocument12 pagesUrinary Tract Infection Anatomy and PhysiologyPaul Anthony Centeno PimentelNo ratings yet
- EpitheliumDocument11 pagesEpitheliumapi-159564163No ratings yet
- Gastro TerminadoDocument6 pagesGastro TerminadoAgus Corazon Barra RamiNo ratings yet
- TomdDocument15 pagesTomdduinnemerriedithcastroNo ratings yet
- Elementary CanalDocument56 pagesElementary Canalmuhammad ijazNo ratings yet
- Digestion and AbsorptionDocument7 pagesDigestion and AbsorptionlalitNo ratings yet
- Digestive System of Human (Biology)Document35 pagesDigestive System of Human (Biology)liofve X viNo ratings yet
- 26 Structure Life History Pila GlobosaDocument8 pages26 Structure Life History Pila Globosafathea111115No ratings yet
- The Structure of The Human Alimentary Canal To Their FunctionsDocument3 pagesThe Structure of The Human Alimentary Canal To Their FunctionsKyng GamariNo ratings yet
- In A Nutshell... : Biology 12 - The Digestive System - Chapter NotesDocument10 pagesIn A Nutshell... : Biology 12 - The Digestive System - Chapter NotesE ZeeNo ratings yet
- Digestion and AbsorptionDocument6 pagesDigestion and AbsorptionMahafuj DudhaknojNo ratings yet
- MG I S9 - The Digestive SystemDocument4 pagesMG I S9 - The Digestive SystemAndrei CristescuNo ratings yet
- Flow and Parts of The Digestive SystemDocument52 pagesFlow and Parts of The Digestive SystemKAnn RuedasNo ratings yet
- Physiology: Anatomy of The Digestive System IntroductionDocument10 pagesPhysiology: Anatomy of The Digestive System IntroductionkyirlessnessNo ratings yet
- Basic Human Anatomy: Lesson 6: The Human Digestive SystemDocument10 pagesBasic Human Anatomy: Lesson 6: The Human Digestive SystemarunNo ratings yet
- Digestive System Anatomy 1 1Document17 pagesDigestive System Anatomy 1 1jsrz9vmj5jNo ratings yet
- A & PDocument2 pagesA & PSahil KulkarniNo ratings yet
- Chapter - 3: Digestive SystemDocument12 pagesChapter - 3: Digestive Systemanantchaurasia11No ratings yet
- Digestive System BioDocument40 pagesDigestive System BioUmm ArafaNo ratings yet
- Digestive LectureDocument20 pagesDigestive LectureKyla Mae ZabalaNo ratings yet
- AnaPhy - Digestive SystemDocument5 pagesAnaPhy - Digestive SystemJan Mark SotoNo ratings yet
- Digestive System and NutritionDocument6 pagesDigestive System and NutritionAthena HuynhNo ratings yet
- The Human Digestive SystemDocument25 pagesThe Human Digestive SystemKrizza Kyle Sarmon MancaoNo ratings yet
- Digestive System: Reported By: Debbie Arboleda Cecille Toquero Joshua Duropan Erika PizarroDocument77 pagesDigestive System: Reported By: Debbie Arboleda Cecille Toquero Joshua Duropan Erika PizarroJezreel OrquinaNo ratings yet
- What Is The Alimentary Canal NOTESSDocument12 pagesWhat Is The Alimentary Canal NOTESSTHAMSANQA SIBANDANo ratings yet
- GuionesDocument7 pagesGuionesEla AnlyNo ratings yet
- Degestive and Skeletal SystemDocument6 pagesDegestive and Skeletal SystemAsad MehmoodNo ratings yet
- Digestive System Anatomy & HistologyDocument7 pagesDigestive System Anatomy & HistologyAbdallah AlasalNo ratings yet
- Digestion and AbsorptionDocument14 pagesDigestion and AbsorptionAnkur YadavNo ratings yet
- 06 Digestive SystemDocument101 pages06 Digestive SystemcelynjasminegNo ratings yet
- Pila GobosaDocument35 pagesPila GobosaTanu SheteNo ratings yet
- Digestive SystemDocument25 pagesDigestive Systembazaar9No ratings yet
- Midterm Examination-BiochemDocument10 pagesMidterm Examination-BiochemRhizel Marie Cauilan FariñasNo ratings yet
- Detailed Study of FrogDocument20 pagesDetailed Study of FrogM. MuvafficaNo ratings yet
- Digestion in HumansDocument21 pagesDigestion in HumansSanaya BansalNo ratings yet
- DigestiveDocument5 pagesDigestivealayca cabatanaNo ratings yet
- Anaphy AGE CzaDocument18 pagesAnaphy AGE CzaJoanna_Edna_C__6752No ratings yet
- The Human Digestion ProcessDocument3 pagesThe Human Digestion Processipunk zuhriNo ratings yet
- OesophagusDocument21 pagesOesophagusparashakthi PRNo ratings yet
- Digestive SystemDocument67 pagesDigestive SystemShacquille Suelto Bautista0% (1)
- Explain The Role of The Following Organs in The Digestive System The Mouth and Salivary GlandsDocument6 pagesExplain The Role of The Following Organs in The Digestive System The Mouth and Salivary GlandsCameronweathersNo ratings yet
- James' Dissection NotesDocument6 pagesJames' Dissection NotesSamuel NyanzuNo ratings yet
- Digestive SystemDocument32 pagesDigestive SystemHashley CastellyNo ratings yet
- Essay # 4. DigeDocument9 pagesEssay # 4. DigeAbdul Karim BurkiNo ratings yet
- Human Digestive System Biology For NEET 2019Document4 pagesHuman Digestive System Biology For NEET 2019misostudyNo ratings yet
- Digestive SystemDocument11 pagesDigestive SystemHaider CheemaNo ratings yet
- To Study The External Features and Digestive System of CockroachesDocument28 pagesTo Study The External Features and Digestive System of CockroachesMinish KatkarNo ratings yet
- The Mouth Cavity, Pharynx, Esophagus and StomachDocument2 pagesThe Mouth Cavity, Pharynx, Esophagus and Stomachகாட்டு வாசிNo ratings yet
- Lamprey DissectionDocument7 pagesLamprey DissectionAnnas KurniawanNo ratings yet
- Digestive System Anatomy PhysiologyDocument20 pagesDigestive System Anatomy PhysiologyKids JangNo ratings yet
- Microsoft PowerPoint - Digestive 2011.Document50 pagesMicrosoft PowerPoint - Digestive 2011.LadyseptianiNo ratings yet
- Anatomy of The Digestive SystemDocument39 pagesAnatomy of The Digestive SystemAngel Ann Gertos Inojales INo ratings yet
- Handout in Science 8: Biology LESSON 1: Organs of The Digestive SystemDocument5 pagesHandout in Science 8: Biology LESSON 1: Organs of The Digestive SystemJanna SarmientoNo ratings yet
- Karthik Bharadwaj'S: PresentationDocument32 pagesKarthik Bharadwaj'S: PresentationMahir AfranNo ratings yet
- Digestive System Anatomy and Physiology - NurseslabsDocument33 pagesDigestive System Anatomy and Physiology - NurseslabsMari FeNo ratings yet
- Digestive System NewDocument26 pagesDigestive System NewJonathan Farne CorpuzNo ratings yet
- Definition of TermsDocument3 pagesDefinition of TermsJoanne Sianson BucayanNo ratings yet
- Nutrition in AnimalsDocument21 pagesNutrition in AnimalsAdityaNo ratings yet
- Digestion and Absorption - NotesDocument15 pagesDigestion and Absorption - NotesSaloni GuptaNo ratings yet
- A Guide for the Dissection of the Dogfish (Squalus Acanthias)From EverandA Guide for the Dissection of the Dogfish (Squalus Acanthias)No ratings yet
- Muscle and Nervous TissueDocument5 pagesMuscle and Nervous Tissueapi-159564163No ratings yet
- Deductive Reasoning TestDocument3 pagesDeductive Reasoning Testapi-159564163No ratings yet
- Sequoia 2nd Class Flyer 2019Document1 pageSequoia 2nd Class Flyer 2019api-159564163No ratings yet
- Bay Area Amphibians15Document59 pagesBay Area Amphibians15api-159564163No ratings yet
- Papillionidae 1Document19 pagesPapillionidae 1api-159564163No ratings yet
- Morganterritory 2aDocument192 pagesMorganterritory 2aapi-159564163No ratings yet
- 12 Making Sense of ItaDocument50 pages12 Making Sense of Itaapi-159564163No ratings yet
- 27 To Pee The UrinarysystemabDocument38 pages27 To Pee The Urinarysystemabapi-159564163No ratings yet
- Project Endocrine Group C - Final 10-14-14Document24 pagesProject Endocrine Group C - Final 10-14-14api-159564163No ratings yet
- Which Is The Optimal Exercise To Strengthen Supraspinatus?: Clinical SciencesDocument5 pagesWhich Is The Optimal Exercise To Strengthen Supraspinatus?: Clinical SciencesVicky PalmaNo ratings yet
- Neuroconducciones Y Electromiografia de Cuatro ExtremidadesDocument6 pagesNeuroconducciones Y Electromiografia de Cuatro ExtremidadesLEON MANUEL JIMENEZ CASTELLANOSNo ratings yet
- 5 Simple Exercises That Will Transform Your Body in Just Four WeeksDocument4 pages5 Simple Exercises That Will Transform Your Body in Just Four WeeksMaria MeșinăNo ratings yet
- Target MRB Test Series 1Document1 pageTarget MRB Test Series 1SanthiyaNo ratings yet
- !adonis Vernalis ProvingDocument17 pages!adonis Vernalis Provingparents021No ratings yet
- Classification of MalocclusionsDocument58 pagesClassification of MalocclusionsKirvase DanNo ratings yet
- Removable Myofunctional Appliance PartialDocument132 pagesRemovable Myofunctional Appliance Partialdr_nilofervevai2360100% (3)
- The Tight Tan Slacks of Dezso Ban - A Back Building Program - Anthony DitilloDocument5 pagesThe Tight Tan Slacks of Dezso Ban - A Back Building Program - Anthony DitilloCD KHUTIYALENo ratings yet
- Biomechnics Mids NotsDocument10 pagesBiomechnics Mids Notstalha.k.rajpootNo ratings yet
- Physiology vs. Anatomy: by Sarah C.P. Willaims Medically Reviewed by Minesh Khatri, MD On July 28, 2021Document2 pagesPhysiology vs. Anatomy: by Sarah C.P. Willaims Medically Reviewed by Minesh Khatri, MD On July 28, 2021exo oxeNo ratings yet
- Primitive Reflexes 240130 213501Document11 pagesPrimitive Reflexes 240130 213501Atharva PawarNo ratings yet
- Worksheet On Brain and Spinal CordDocument13 pagesWorksheet On Brain and Spinal Cordcris baligodNo ratings yet
- LSM Grade 3 Science 1st Trim Exam SY 2009-2010 Answer KeyDocument8 pagesLSM Grade 3 Science 1st Trim Exam SY 2009-2010 Answer KeyMauie Flores100% (2)
- AnaPhy SAS 1Document5 pagesAnaPhy SAS 1Zylith NanaseNo ratings yet
- UROGYNECOLOGY: Urinary Incontinence and Modalities of Treatment by Prof - Dr.Vesna AntovskaDocument19 pagesUROGYNECOLOGY: Urinary Incontinence and Modalities of Treatment by Prof - Dr.Vesna AntovskaVesna Antovska100% (1)
- Bodyweight Fitness Progressions - Version 5.4Document1 pageBodyweight Fitness Progressions - Version 5.4Spinelli PietroNo ratings yet
- Iliac Crest Grafting For Mandibular ReconstructionDocument10 pagesIliac Crest Grafting For Mandibular Reconstructiontanvi vijayNo ratings yet
- Cranial Nerves - MRI Anatomy and Cases: A Santa SibiuDocument121 pagesCranial Nerves - MRI Anatomy and Cases: A Santa SibiuAndreea CimpoiNo ratings yet
- Lesson 12 HeartDocument9 pagesLesson 12 HeartEly FructuosoNo ratings yet
- HeadDocument360 pagesHeadfeliciostenio100% (1)
- Cardiovascular SystemDocument10 pagesCardiovascular SystemMr. Christian ParabuacNo ratings yet
- Cervical GoniometeryDocument39 pagesCervical GoniometerySimranjit KaurNo ratings yet
- Placenta Prayer of ReleaseDocument4 pagesPlacenta Prayer of Releaselizette FourieNo ratings yet
- Human Body WebQuestDocument6 pagesHuman Body WebQuestjennyferchoNo ratings yet
- TracheostomyDocument28 pagesTracheostomyprivatejarjarbinksNo ratings yet
- Autonomic Nervous SystemDocument40 pagesAutonomic Nervous SystemmarciaNo ratings yet
- Lecture 3Document25 pagesLecture 3Yoshe RionandoNo ratings yet
- Muscles of The ShoulderDocument79 pagesMuscles of The Shoulderali.algazali20No ratings yet