
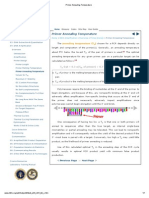
Primer Design: (In This Case The Word Primer Means An Oligonucleotide)
Primer Design: (In This Case The Word Primer Means An Oligonucleotide)
You might also like
- PCR Amplification Lab ReportDocument5 pagesPCR Amplification Lab ReportWilson Chan100% (7)
- Maria Afzal M.phill-03 Submitted To: Prof. Dr. Farkhanda JabeenDocument44 pagesMaria Afzal M.phill-03 Submitted To: Prof. Dr. Farkhanda JabeenMuhammad MushtaqNo ratings yet
- Primer Design 2013 PDFDocument58 pagesPrimer Design 2013 PDFMaila EscuderoNo ratings yet
- Chapter8 PrimerdesigningDocument8 pagesChapter8 PrimerdesigningSakshi SangleNo ratings yet
- Primer Design ExerciseDocument34 pagesPrimer Design ExerciseRita karen Pacheco CabañasNo ratings yet
- How To Design PrimersDocument14 pagesHow To Design PrimersDeepak RawoolNo ratings yet
- Primer Designing For PCR: ColloquiumDocument3 pagesPrimer Designing For PCR: ColloquiumSakshi SangleNo ratings yet
- PrimersDocument2 pagesPrimersmattiullahkhan860No ratings yet
- Principle of The PCR: This Technique Is Invented in 1984 by Kary Mulis. The Purpose of A PCRDocument11 pagesPrinciple of The PCR: This Technique Is Invented in 1984 by Kary Mulis. The Purpose of A PCRantesar reheemNo ratings yet
- Taqman Probe Design Guidelines - May 2018Document5 pagesTaqman Probe Design Guidelines - May 2018AviNo ratings yet
- Primer Deigning 1Document18 pagesPrimer Deigning 1Pakistan ZindaabdaNo ratings yet
- Prerequisite of A PCR: Logarithmic and Controlled FashionDocument18 pagesPrerequisite of A PCR: Logarithmic and Controlled FashionlkokodkodNo ratings yet
- 4.PCR Primer Designing (Deepa Chouhan) - 1Document14 pages4.PCR Primer Designing (Deepa Chouhan) - 1VK 360 CREATIONSNo ratings yet
- General PCR Guidelines: PrimersDocument13 pagesGeneral PCR Guidelines: PrimersAdi NugrahaNo ratings yet
- Primer Design For PCRDocument1 pagePrimer Design For PCRdoubleyouem2003No ratings yet
- PCRDocument11 pagesPCRpkhkhawar100% (1)
- PCR Primer DesignDocument13 pagesPCR Primer DesignAiman ArshadNo ratings yet
- Polymerase Chain Reaction ObjectivesDocument25 pagesPolymerase Chain Reaction ObjectivestinasheNo ratings yet
- Primer Designing For PCRDocument13 pagesPrimer Designing For PCRiftikharsubha5No ratings yet
- Jenis-Jenis PCRDocument6 pagesJenis-Jenis PCRDwi Novia PutriNo ratings yet
- Chapter-6 The Polymerase Chain Reaction: MBO 415-Diagnostic Microbiology Lecture-10 Ahnaf Tahmid SaqifDocument20 pagesChapter-6 The Polymerase Chain Reaction: MBO 415-Diagnostic Microbiology Lecture-10 Ahnaf Tahmid SaqifASADULLAH AL GALIBNo ratings yet
- PCR Primer Design PDFDocument62 pagesPCR Primer Design PDFnesariaNo ratings yet
- Adv 4.Document18 pagesAdv 4.harseen rahimNo ratings yet
- PCR An OverviewDocument41 pagesPCR An OverviewnassradabourNo ratings yet
- The Polymerase Chain Reaction and Sequence-Tagged SitesDocument7 pagesThe Polymerase Chain Reaction and Sequence-Tagged SitesPisi MicaNo ratings yet
- Please Complete The .: Workshop Sign-In FormDocument18 pagesPlease Complete The .: Workshop Sign-In FormamiraNo ratings yet
- Tutorial 6 Molecular TechniquesDocument4 pagesTutorial 6 Molecular TechniquesJason LeeNo ratings yet
- Nucleic Acid Amplification PDFDocument9 pagesNucleic Acid Amplification PDFSajeebChandraNo ratings yet
- Polymerase Chain ReactionDocument27 pagesPolymerase Chain ReactionundalliNo ratings yet
- Bauer Core Real Time GuidelinesDocument5 pagesBauer Core Real Time GuidelinesISmi MumtaZahNo ratings yet
- Polymerase Chain Reaction (PCR)Document37 pagesPolymerase Chain Reaction (PCR)Haleema SultanNo ratings yet
- Aspect On PCR ReactionDocument45 pagesAspect On PCR ReactionsunfonemmaNo ratings yet
- Polymerase Chain ReactionDocument16 pagesPolymerase Chain ReactionAbdulati Abu RewillaNo ratings yet
- Primer Designing2Document5 pagesPrimer Designing2Angel ZhangNo ratings yet
- PCR Basics: Kanadi SumaprajaDocument31 pagesPCR Basics: Kanadi SumaprajaSamrichardNo ratings yet
- Polymerase Chain ReactionDocument60 pagesPolymerase Chain Reactionclarice_anneNo ratings yet
- PCR1Document4 pagesPCR1Hina NoorNo ratings yet
- Nucleic Acid Manipulation: The Polymerase Chain ReactionDocument22 pagesNucleic Acid Manipulation: The Polymerase Chain Reactionamor_panopioNo ratings yet
- Addgene: Protocol - How To Design PrimersDocument1 pageAddgene: Protocol - How To Design PrimersArantxa Ortega LeonNo ratings yet
- 2019-20 Bioinformatics CDocument28 pages2019-20 Bioinformatics CbeaNo ratings yet
- Continuous Assessment: Concentration's Increment in Case of Insert and LigaseDocument8 pagesContinuous Assessment: Concentration's Increment in Case of Insert and LigaseDebapriya HazraNo ratings yet
- LECT-12 The Polymerase Chain Reaction (20192020-1)Document23 pagesLECT-12 The Polymerase Chain Reaction (20192020-1)Ikrimah FithriyandiniNo ratings yet
- Primer Annealing TemperatureDocument1 pagePrimer Annealing TemperatureManu Mallahalli ShanthappaNo ratings yet
- Chapter 5 - PrimerDesign and Restriction Digestion Analysis PRINTDocument112 pagesChapter 5 - PrimerDesign and Restriction Digestion Analysis PRINTshuma chemedaNo ratings yet
- PCR PresentationDocument34 pagesPCR Presentationdurgaprasadhembram1999No ratings yet
- Polymerase Chain ReactionDocument47 pagesPolymerase Chain ReactionshvnagaNo ratings yet
- Lecture 4: Detecting Mutations and Mutation Consequences Learning GoalsDocument11 pagesLecture 4: Detecting Mutations and Mutation Consequences Learning GoalsAngelica SmithNo ratings yet
- Polymerase Chain Reaction (PCR)Document3 pagesPolymerase Chain Reaction (PCR)biotech_vidhyaNo ratings yet
- pcr شرح مفصل عنDocument26 pagespcr شرح مفصل عنZainab HasanNo ratings yet
- Lorenz2012 PDFDocument15 pagesLorenz2012 PDFCatherine Cinthya RIMAC PANEZNo ratings yet
- × Aboratory Tools and TechniquesDocument23 pages× Aboratory Tools and TechniquesjoshigautaNo ratings yet
- PCR Primer Design 2013Document29 pagesPCR Primer Design 2013Kartikeya SinghNo ratings yet
- DNA Techniques (PCR and Sequencing)Document4 pagesDNA Techniques (PCR and Sequencing)ffarrugia12No ratings yet
- The Olymerase Hain Eaction: Zhang-Haifeng Department of BiochemistryDocument42 pagesThe Olymerase Hain Eaction: Zhang-Haifeng Department of Biochemistryapi-19916399No ratings yet
- Next Generation Sequencing and Sequence Assembly: Methodologies and AlgorithmsFrom EverandNext Generation Sequencing and Sequence Assembly: Methodologies and AlgorithmsNo ratings yet
- DNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensFrom EverandDNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensNo ratings yet
- Nuclear Medicine Technology: Review Questions for the Board ExaminationsFrom EverandNuclear Medicine Technology: Review Questions for the Board ExaminationsNo ratings yet
- Biophysical Analysis of Membrane Proteins: Investigating Structure and FunctionFrom EverandBiophysical Analysis of Membrane Proteins: Investigating Structure and FunctionNo ratings yet



























































Download as pdf or txt
You might also like
- PCR Amplification Lab ReportDocument5 pagesPCR Amplification Lab ReportWilson Chan100% (7)
- Maria Afzal M.phill-03 Submitted To: Prof. Dr. Farkhanda JabeenDocument44 pagesMaria Afzal M.phill-03 Submitted To: Prof. Dr. Farkhanda JabeenMuhammad MushtaqNo ratings yet
- Primer Design 2013 PDFDocument58 pagesPrimer Design 2013 PDFMaila EscuderoNo ratings yet
- Chapter8 PrimerdesigningDocument8 pagesChapter8 PrimerdesigningSakshi SangleNo ratings yet
- Primer Design ExerciseDocument34 pagesPrimer Design ExerciseRita karen Pacheco CabañasNo ratings yet
- How To Design PrimersDocument14 pagesHow To Design PrimersDeepak RawoolNo ratings yet
- Primer Designing For PCR: ColloquiumDocument3 pagesPrimer Designing For PCR: ColloquiumSakshi SangleNo ratings yet
- PrimersDocument2 pagesPrimersmattiullahkhan860No ratings yet
- Principle of The PCR: This Technique Is Invented in 1984 by Kary Mulis. The Purpose of A PCRDocument11 pagesPrinciple of The PCR: This Technique Is Invented in 1984 by Kary Mulis. The Purpose of A PCRantesar reheemNo ratings yet
- Taqman Probe Design Guidelines - May 2018Document5 pagesTaqman Probe Design Guidelines - May 2018AviNo ratings yet
- Primer Deigning 1Document18 pagesPrimer Deigning 1Pakistan ZindaabdaNo ratings yet
- Prerequisite of A PCR: Logarithmic and Controlled FashionDocument18 pagesPrerequisite of A PCR: Logarithmic and Controlled FashionlkokodkodNo ratings yet
- 4.PCR Primer Designing (Deepa Chouhan) - 1Document14 pages4.PCR Primer Designing (Deepa Chouhan) - 1VK 360 CREATIONSNo ratings yet
- General PCR Guidelines: PrimersDocument13 pagesGeneral PCR Guidelines: PrimersAdi NugrahaNo ratings yet
- Primer Design For PCRDocument1 pagePrimer Design For PCRdoubleyouem2003No ratings yet
- PCRDocument11 pagesPCRpkhkhawar100% (1)
- PCR Primer DesignDocument13 pagesPCR Primer DesignAiman ArshadNo ratings yet
- Polymerase Chain Reaction ObjectivesDocument25 pagesPolymerase Chain Reaction ObjectivestinasheNo ratings yet
- Primer Designing For PCRDocument13 pagesPrimer Designing For PCRiftikharsubha5No ratings yet
- Jenis-Jenis PCRDocument6 pagesJenis-Jenis PCRDwi Novia PutriNo ratings yet
- Chapter-6 The Polymerase Chain Reaction: MBO 415-Diagnostic Microbiology Lecture-10 Ahnaf Tahmid SaqifDocument20 pagesChapter-6 The Polymerase Chain Reaction: MBO 415-Diagnostic Microbiology Lecture-10 Ahnaf Tahmid SaqifASADULLAH AL GALIBNo ratings yet
- PCR Primer Design PDFDocument62 pagesPCR Primer Design PDFnesariaNo ratings yet
- Adv 4.Document18 pagesAdv 4.harseen rahimNo ratings yet
- PCR An OverviewDocument41 pagesPCR An OverviewnassradabourNo ratings yet
- The Polymerase Chain Reaction and Sequence-Tagged SitesDocument7 pagesThe Polymerase Chain Reaction and Sequence-Tagged SitesPisi MicaNo ratings yet
- Please Complete The .: Workshop Sign-In FormDocument18 pagesPlease Complete The .: Workshop Sign-In FormamiraNo ratings yet
- Tutorial 6 Molecular TechniquesDocument4 pagesTutorial 6 Molecular TechniquesJason LeeNo ratings yet
- Nucleic Acid Amplification PDFDocument9 pagesNucleic Acid Amplification PDFSajeebChandraNo ratings yet
- Polymerase Chain ReactionDocument27 pagesPolymerase Chain ReactionundalliNo ratings yet
- Bauer Core Real Time GuidelinesDocument5 pagesBauer Core Real Time GuidelinesISmi MumtaZahNo ratings yet
- Polymerase Chain Reaction (PCR)Document37 pagesPolymerase Chain Reaction (PCR)Haleema SultanNo ratings yet
- Aspect On PCR ReactionDocument45 pagesAspect On PCR ReactionsunfonemmaNo ratings yet
- Polymerase Chain ReactionDocument16 pagesPolymerase Chain ReactionAbdulati Abu RewillaNo ratings yet
- Primer Designing2Document5 pagesPrimer Designing2Angel ZhangNo ratings yet
- PCR Basics: Kanadi SumaprajaDocument31 pagesPCR Basics: Kanadi SumaprajaSamrichardNo ratings yet
- Polymerase Chain ReactionDocument60 pagesPolymerase Chain Reactionclarice_anneNo ratings yet
- PCR1Document4 pagesPCR1Hina NoorNo ratings yet
- Nucleic Acid Manipulation: The Polymerase Chain ReactionDocument22 pagesNucleic Acid Manipulation: The Polymerase Chain Reactionamor_panopioNo ratings yet
- Addgene: Protocol - How To Design PrimersDocument1 pageAddgene: Protocol - How To Design PrimersArantxa Ortega LeonNo ratings yet
- 2019-20 Bioinformatics CDocument28 pages2019-20 Bioinformatics CbeaNo ratings yet
- Continuous Assessment: Concentration's Increment in Case of Insert and LigaseDocument8 pagesContinuous Assessment: Concentration's Increment in Case of Insert and LigaseDebapriya HazraNo ratings yet
- LECT-12 The Polymerase Chain Reaction (20192020-1)Document23 pagesLECT-12 The Polymerase Chain Reaction (20192020-1)Ikrimah FithriyandiniNo ratings yet
- Primer Annealing TemperatureDocument1 pagePrimer Annealing TemperatureManu Mallahalli ShanthappaNo ratings yet
- Chapter 5 - PrimerDesign and Restriction Digestion Analysis PRINTDocument112 pagesChapter 5 - PrimerDesign and Restriction Digestion Analysis PRINTshuma chemedaNo ratings yet
- PCR PresentationDocument34 pagesPCR Presentationdurgaprasadhembram1999No ratings yet
- Polymerase Chain ReactionDocument47 pagesPolymerase Chain ReactionshvnagaNo ratings yet
- Lecture 4: Detecting Mutations and Mutation Consequences Learning GoalsDocument11 pagesLecture 4: Detecting Mutations and Mutation Consequences Learning GoalsAngelica SmithNo ratings yet
- Polymerase Chain Reaction (PCR)Document3 pagesPolymerase Chain Reaction (PCR)biotech_vidhyaNo ratings yet
- pcr شرح مفصل عنDocument26 pagespcr شرح مفصل عنZainab HasanNo ratings yet
- Lorenz2012 PDFDocument15 pagesLorenz2012 PDFCatherine Cinthya RIMAC PANEZNo ratings yet
- × Aboratory Tools and TechniquesDocument23 pages× Aboratory Tools and TechniquesjoshigautaNo ratings yet
- PCR Primer Design 2013Document29 pagesPCR Primer Design 2013Kartikeya SinghNo ratings yet
- DNA Techniques (PCR and Sequencing)Document4 pagesDNA Techniques (PCR and Sequencing)ffarrugia12No ratings yet
- The Olymerase Hain Eaction: Zhang-Haifeng Department of BiochemistryDocument42 pagesThe Olymerase Hain Eaction: Zhang-Haifeng Department of Biochemistryapi-19916399No ratings yet
- Next Generation Sequencing and Sequence Assembly: Methodologies and AlgorithmsFrom EverandNext Generation Sequencing and Sequence Assembly: Methodologies and AlgorithmsNo ratings yet
- DNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensFrom EverandDNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensNo ratings yet
- Nuclear Medicine Technology: Review Questions for the Board ExaminationsFrom EverandNuclear Medicine Technology: Review Questions for the Board ExaminationsNo ratings yet
- Biophysical Analysis of Membrane Proteins: Investigating Structure and FunctionFrom EverandBiophysical Analysis of Membrane Proteins: Investigating Structure and FunctionNo ratings yet