Download as pdf or txt
You might also like
- Humana Genetika - RepetitorijumDocument72 pagesHumana Genetika - RepetitorijumТамара Васић95% (22)
- Humana Genetika Ispitna Pitanja I Odgovori Za Studente MedicineDocument82 pagesHumana Genetika Ispitna Pitanja I Odgovori Za Studente MedicineOgnjen Milovic100% (3)
- Nukleinske Kiseline Ppt1Document14 pagesNukleinske Kiseline Ppt1Edin VišnjićNo ratings yet
- Nukleinske KiselineDocument93 pagesNukleinske KiselineRastko ZečevićNo ratings yet
- Struktura I Hem Osob DNA PDFDocument76 pagesStruktura I Hem Osob DNA PDFdženanam_2No ratings yet
- Celokupno Gradivo Za Integralni Test PDFDocument17 pagesCelokupno Gradivo Za Integralni Test PDFBosko JovancevicNo ratings yet
- Sličnosti I Razlike DNK I RNK - Batilovic DamirDocument18 pagesSličnosti I Razlike DNK I RNK - Batilovic Damirakademac groupNo ratings yet
- Molekularna Biologija I DNK (Compatibility Mode)Document20 pagesMolekularna Biologija I DNK (Compatibility Mode)Sonja TodoricNo ratings yet
- Glavna SkriptaDocument83 pagesGlavna SkriptaMohanad YoByNo ratings yet
- Humana GenetikaDocument44 pagesHumana GenetikaNemanja Stanisavljevic100% (1)
- Vežba 3 - Tehnike U Molekularnoj BiologijiDocument30 pagesVežba 3 - Tehnike U Molekularnoj BiologijiAnonymous uqG689HNo ratings yet
- Nukleinske KiselineDocument6 pagesNukleinske Kiselinestasa.miletic.2020.8No ratings yet
- Biokemija Proteina I Nukleinskih Kiselina - PredavanjaDocument40 pagesBiokemija Proteina I Nukleinskih Kiselina - PredavanjaElvir M100% (1)
- Nukleotidi I Nukleinske KiselineDocument30 pagesNukleotidi I Nukleinske KiselineAdina AhmetovicNo ratings yet
- Replikacija DNKDocument10 pagesReplikacija DNKRadica MiljkovicNo ratings yet
- I Osnovi Molekularne Biologije ProkariotaDocument79 pagesI Osnovi Molekularne Biologije ProkariotaMilenaNo ratings yet
- RNA I TranskripcijaDocument40 pagesRNA I TranskripcijaMiki MausNo ratings yet
- TranslacijaDocument42 pagesTranslacijaAnaNikolov100% (1)
- ReplikacijaDocument2 pagesReplikacijaSara Azra DautovićNo ratings yet
- Lekcija - Nukleinske Kiseline - Za Sajt FrizeriDocument3 pagesLekcija - Nukleinske Kiseline - Za Sajt Frizeriapi-270998884No ratings yet
- Osnovi Molekularne Biologije ProkariotaDocument79 pagesOsnovi Molekularne Biologije ProkariotaEdin SmajicNo ratings yet
- Матурски рад РНК и транскрипцијаDocument12 pagesМатурски рад РНК и транскрипцијаSenkaNo ratings yet
- TJ 12 Metode Oksido-Redukcijskih TitracijaDocument44 pagesTJ 12 Metode Oksido-Redukcijskih TitracijaGorannnNo ratings yet
- Нуклеинске киселинеDocument14 pagesНуклеинске киселинеDusan CavicNo ratings yet
- Obrada RNKDocument57 pagesObrada RNKNenad ĐorđevićNo ratings yet
- Pojam RNKDocument14 pagesPojam RNKIgor ĐorđevićNo ratings yet
- Нуклеинске Киселине 1 1Document11 pagesНуклеинске Киселине 1 1Milica StojmenovicNo ratings yet
- Основна Догма Молекуларне БиологијеDocument19 pagesОсновна Догма Молекуларне БиологијеСнежана СтикићNo ratings yet
- Nukleinske KiselineDocument16 pagesNukleinske KiselineЛука ТрбовићNo ratings yet
- Nukleinske Kiseline TanjaDocument58 pagesNukleinske Kiseline TanjaMilan BoskovicNo ratings yet
- Uloga RNK U Katalitičkim ProcesimaDocument3 pagesUloga RNK U Katalitičkim ProcesimaVladimir VelimirovicNo ratings yet
- 4 Celija I Biomolekuli III Nukleinske Kiseline I Sinteza Proteina PDFDocument47 pages4 Celija I Biomolekuli III Nukleinske Kiseline I Sinteza Proteina PDFMirjana MihajlovićNo ratings yet
- Dezoksiribonukleinska KiselinaDocument2 pagesDezoksiribonukleinska KiselinaNikola LazarevicNo ratings yet
- РЕПЛИКАЦИЈАDocument4 pagesРЕПЛИКАЦИЈАAndrea KovačevićNo ratings yet
- REPLIKACIJADocument5 pagesREPLIKACIJAMaja BorkovićNo ratings yet
- Molekularna GenetikaDocument39 pagesMolekularna GenetikaNikolić MladenNo ratings yet
- RNKDocument9 pagesRNKFilip SrdijaNo ratings yet
- Helati HemijaDocument5 pagesHelati HemijaSelmir IsakovićNo ratings yet
- Skripta GenetikaDocument24 pagesSkripta GenetikamikiksenijaNo ratings yet
- ĆelijaDocument8 pagesĆelijaKatarina MiticNo ratings yet
- Нуклеинске киселинеDocument15 pagesНуклеинске киселинеKristinaNo ratings yet
- Dinamička Organizacija ĆelijeDocument6 pagesDinamička Organizacija ĆelijeTamaraŠkrbićNo ratings yet
- EnzimDocument14 pagesEnzimmediteraneo011No ratings yet
- Mitohondrijska DNKDocument5 pagesMitohondrijska DNKHexaGlobinNo ratings yet
- Vpred-IIsem 09Document82 pagesVpred-IIsem 09Visnja VisnjicNo ratings yet
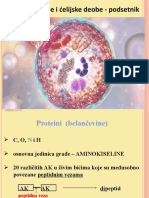
- Biologija Ćelije I Ćelijske Deobe - Podsetnik Za UčenikeDocument17 pagesBiologija Ćelije I Ćelijske Deobe - Podsetnik Za UčenikestefanaNo ratings yet
- МОЛЕКУЛАРНА БИОЛОГИЈАDocument12 pagesМОЛЕКУЛАРНА БИОЛОГИЈАBogdan AleksićNo ratings yet
- DNK I RNKDocument13 pagesDNK I RNKAna VelasevicNo ratings yet
- 4 RastvaranjeUvodi Hidratacija AkvaKompleksiDocument15 pages4 RastvaranjeUvodi Hidratacija AkvaKompleksiMilica DjericNo ratings yet
- Biologija Građa BićaDocument8 pagesBiologija Građa BićaLazar MedarNo ratings yet
- Nukleinske Kis1Document15 pagesNukleinske Kis1Tatjana RaičevićNo ratings yet
- ДНК И РНКDocument12 pagesДНК И РНКTamara NikolićNo ratings yet