
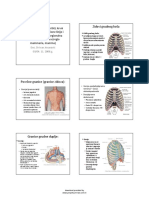
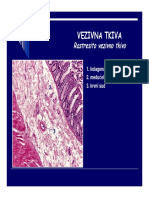
05 Vrste Odbacivanja Organa
05 Vrste Odbacivanja Organa
You might also like
- Opisite Faze Sazrijevanja TDocument2 pagesOpisite Faze Sazrijevanja TTamaraNo ratings yet
- Test Hirurgija - Specijalna PitanjaDocument22 pagesTest Hirurgija - Specijalna PitanjaNenad ĐorđevićNo ratings yet
- Patologija SkraćenoDocument139 pagesPatologija SkraćenoMatej PeretinNo ratings yet
- Mali Mozak Medjumozak ScribdDocument19 pagesMali Mozak Medjumozak ScribdGoran100% (1)
- Mitoza ScribdDocument23 pagesMitoza ScribdGoran100% (1)
- Zavrsni Iz Fiziologije Biljaka 3 2Document12 pagesZavrsni Iz Fiziologije Biljaka 3 2Anonymous bFcaYbRROCNo ratings yet
- PRED 2-ZAPALJENJE (Inflamatio)Document222 pagesPRED 2-ZAPALJENJE (Inflamatio)Jovana JojicNo ratings yet
- GenetikaDocument42 pagesGenetikaAnelaMDNo ratings yet
- Patologija Sa PatofiziologijomDocument120 pagesPatologija Sa PatofiziologijomDragan VeljkovicNo ratings yet
- MEFOS - Pitanja S Kolokvija Iz BiologijeDocument17 pagesMEFOS - Pitanja S Kolokvija Iz BiologijeKrasic IvanNo ratings yet
- Aortna Stenoza - PrezentacijaDocument17 pagesAortna Stenoza - Prezentacijaabi1607No ratings yet
- Fiziologija Nadbubrezne ZlijezdeDocument9 pagesFiziologija Nadbubrezne Zlijezdeammar_okanovicNo ratings yet
- Vezivno TkivoDocument39 pagesVezivno TkivoBlake201No ratings yet
- Dijagnoza LikvoraDocument14 pagesDijagnoza LikvoraMatija PajićNo ratings yet
- CestodeDocument27 pagesCestodeAdis Šišić100% (1)
- COR AnatomijaDocument25 pagesCOR Anatomijacvjetic cvjeticNo ratings yet
- EKG InterpretacijaDocument3 pagesEKG InterpretacijaJana KupusovicNo ratings yet
- Tumori Koštano Zglobnog Sistema 2012Document35 pagesTumori Koštano Zglobnog Sistema 2012Teodora StevanovicNo ratings yet
- Pregled Nervnog SistemaDocument43 pagesPregled Nervnog SistemaStevan100% (1)
- Naprednato Odrzuvanje Vo Zivot (Advanced Life Support)Document33 pagesNaprednato Odrzuvanje Vo Zivot (Advanced Life Support)bala bat0% (1)
- Fiziologija KVSDocument20 pagesFiziologija KVSMarko GajicNo ratings yet
- MD - Diferencijalna Dijagnoza Sindroma Mlitavog OdojčetaDocument6 pagesMD - Diferencijalna Dijagnoza Sindroma Mlitavog OdojčetaNestor CastilloNo ratings yet
- 2 Ciklus Tehnike UlltrazvukaDocument153 pages2 Ciklus Tehnike UlltrazvukaHanan VrcićNo ratings yet
- Bronhoskopija U JILDocument35 pagesBronhoskopija U JILvladimir pavlovicNo ratings yet
- Hipertenzivna KrizaDocument35 pagesHipertenzivna Krizamaja86.milojevicNo ratings yet
- Oštrina VidaDocument21 pagesOštrina Vidakochab 7No ratings yet
- Anatomija SrcaDocument22 pagesAnatomija SrcaVesna FilipovskiNo ratings yet
- Fizioloske Osnove Psihickih PojavaDocument3 pagesFizioloske Osnove Psihickih PojavaLjubomir KljajicNo ratings yet
- Predavanje - Zidovi Grudnog KosaDocument11 pagesPredavanje - Zidovi Grudnog KosaManda GrgicNo ratings yet
- 22 Patofiziologija Nervnog Sistema PDFDocument30 pages22 Patofiziologija Nervnog Sistema PDFTarikNo ratings yet
- Pitanja Za HordateDocument8 pagesPitanja Za HordateAnonymous bFcaYbRROCNo ratings yet
- Poli TraumaDocument20 pagesPoli TraumaBenjamin MalkicNo ratings yet
- Benigni Tumori Kostiju VilicaDocument32 pagesBenigni Tumori Kostiju VilicaHelena Tomas100% (1)
- Bolesti - Intrakranijalni TumoriDocument29 pagesBolesti - Intrakranijalni Tumorizixdiddy100% (1)
- Centralna SinapsaDocument16 pagesCentralna SinapsabetmenNo ratings yet
- Fiziologija SrcaDocument12 pagesFiziologija SrcaNevena BrankovicNo ratings yet
- PREZENTACIJEDocument6 pagesPREZENTACIJENickNo ratings yet
- Hromozomske AberacijeDocument1 pageHromozomske AberacijeAleksandra PericicNo ratings yet
- Auskultacija SrcaDocument8 pagesAuskultacija SrcaKemko KemsNo ratings yet
- Patologija 5Document95 pagesPatologija 5Jelena Dajana Bandovic PanicNo ratings yet
- Difuzija, Osmoza, OsmometriDocument7 pagesDifuzija, Osmoza, OsmometriMateam55No ratings yet
- Disk-Difuziona Metoda Ispitivanja Osjetljivosti Bakterija Na AntibiotikeDocument11 pagesDisk-Difuziona Metoda Ispitivanja Osjetljivosti Bakterija Na AntibiotikeMustafa Korjenić100% (3)
- Laboratorijske ProcedureDocument24 pagesLaboratorijske ProcedureJasmina Farmasi SamardžićNo ratings yet
- Samir I AnaDocument11 pagesSamir I AnaSamir CehicNo ratings yet
- Rak DojkeDocument8 pagesRak DojkeAhmet SalihovicNo ratings yet
- Koagulacija - Predavanje 2014Document18 pagesKoagulacija - Predavanje 2014vanja24312No ratings yet
- 002 Kranijalni Zivci I Putevi CnsDocument39 pages002 Kranijalni Zivci I Putevi CnsIvana Lukovic50% (2)
- Vezivno TkivoDocument17 pagesVezivno TkivoRadna Skela1No ratings yet
- Voda I Minerali Specijalizanti1Document192 pagesVoda I Minerali Specijalizanti1Dejan Todorovic100% (1)
- 1 - Prijenos Tvari Kroz Staničnu Membranu - HoDocument8 pages1 - Prijenos Tvari Kroz Staničnu Membranu - HoTamara Bluebell ﭢNo ratings yet
- FIA-Uputstvo Za Popunjavanje Uplatnice PDFDocument2 pagesFIA-Uputstvo Za Popunjavanje Uplatnice PDFabaz_zbNo ratings yet
- Bilje ĆelijeDocument4 pagesBilje ĆelijeSelma SakićNo ratings yet
- VEZBA 4 Laboratorijska DG Gljivicnih InfekcijaDocument45 pagesVEZBA 4 Laboratorijska DG Gljivicnih InfekcijaMDNo ratings yet
- Medicinski Znacaj Buke I VibracijaDocument7 pagesMedicinski Znacaj Buke I VibracijaAleksandar Aca DvornickiNo ratings yet
- Kranijalni ZivciDocument20 pagesKranijalni Zivcikarlaj97100% (1)
- Limfociti BDocument6 pagesLimfociti BIva PotočnjakNo ratings yet
- Skripta Imunogenetika Sa Osnovama ImunologijeDocument59 pagesSkripta Imunogenetika Sa Osnovama ImunologijeZerina CamdzicNo ratings yet
- Imunologija PredavanjaDocument268 pagesImunologija PredavanjascepanNo ratings yet
- Imuni Sistem, Ćelije Imunog SistemaDocument30 pagesImuni Sistem, Ćelije Imunog SistemaHany Tasha DexterNo ratings yet
- Citologija I GenetikaDocument49 pagesCitologija I GenetikaMsPaparazzyNo ratings yet






















































Download as doc, pdf, or txt
You might also like
- Opisite Faze Sazrijevanja TDocument2 pagesOpisite Faze Sazrijevanja TTamaraNo ratings yet
- Test Hirurgija - Specijalna PitanjaDocument22 pagesTest Hirurgija - Specijalna PitanjaNenad ĐorđevićNo ratings yet
- Patologija SkraćenoDocument139 pagesPatologija SkraćenoMatej PeretinNo ratings yet
- Mali Mozak Medjumozak ScribdDocument19 pagesMali Mozak Medjumozak ScribdGoran100% (1)
- Mitoza ScribdDocument23 pagesMitoza ScribdGoran100% (1)
- Zavrsni Iz Fiziologije Biljaka 3 2Document12 pagesZavrsni Iz Fiziologije Biljaka 3 2Anonymous bFcaYbRROCNo ratings yet
- PRED 2-ZAPALJENJE (Inflamatio)Document222 pagesPRED 2-ZAPALJENJE (Inflamatio)Jovana JojicNo ratings yet
- GenetikaDocument42 pagesGenetikaAnelaMDNo ratings yet
- Patologija Sa PatofiziologijomDocument120 pagesPatologija Sa PatofiziologijomDragan VeljkovicNo ratings yet
- MEFOS - Pitanja S Kolokvija Iz BiologijeDocument17 pagesMEFOS - Pitanja S Kolokvija Iz BiologijeKrasic IvanNo ratings yet
- Aortna Stenoza - PrezentacijaDocument17 pagesAortna Stenoza - Prezentacijaabi1607No ratings yet
- Fiziologija Nadbubrezne ZlijezdeDocument9 pagesFiziologija Nadbubrezne Zlijezdeammar_okanovicNo ratings yet
- Vezivno TkivoDocument39 pagesVezivno TkivoBlake201No ratings yet
- Dijagnoza LikvoraDocument14 pagesDijagnoza LikvoraMatija PajićNo ratings yet
- CestodeDocument27 pagesCestodeAdis Šišić100% (1)
- COR AnatomijaDocument25 pagesCOR Anatomijacvjetic cvjeticNo ratings yet
- EKG InterpretacijaDocument3 pagesEKG InterpretacijaJana KupusovicNo ratings yet
- Tumori Koštano Zglobnog Sistema 2012Document35 pagesTumori Koštano Zglobnog Sistema 2012Teodora StevanovicNo ratings yet
- Pregled Nervnog SistemaDocument43 pagesPregled Nervnog SistemaStevan100% (1)
- Naprednato Odrzuvanje Vo Zivot (Advanced Life Support)Document33 pagesNaprednato Odrzuvanje Vo Zivot (Advanced Life Support)bala bat0% (1)
- Fiziologija KVSDocument20 pagesFiziologija KVSMarko GajicNo ratings yet
- MD - Diferencijalna Dijagnoza Sindroma Mlitavog OdojčetaDocument6 pagesMD - Diferencijalna Dijagnoza Sindroma Mlitavog OdojčetaNestor CastilloNo ratings yet
- 2 Ciklus Tehnike UlltrazvukaDocument153 pages2 Ciklus Tehnike UlltrazvukaHanan VrcićNo ratings yet
- Bronhoskopija U JILDocument35 pagesBronhoskopija U JILvladimir pavlovicNo ratings yet
- Hipertenzivna KrizaDocument35 pagesHipertenzivna Krizamaja86.milojevicNo ratings yet
- Oštrina VidaDocument21 pagesOštrina Vidakochab 7No ratings yet
- Anatomija SrcaDocument22 pagesAnatomija SrcaVesna FilipovskiNo ratings yet
- Fizioloske Osnove Psihickih PojavaDocument3 pagesFizioloske Osnove Psihickih PojavaLjubomir KljajicNo ratings yet
- Predavanje - Zidovi Grudnog KosaDocument11 pagesPredavanje - Zidovi Grudnog KosaManda GrgicNo ratings yet
- 22 Patofiziologija Nervnog Sistema PDFDocument30 pages22 Patofiziologija Nervnog Sistema PDFTarikNo ratings yet
- Pitanja Za HordateDocument8 pagesPitanja Za HordateAnonymous bFcaYbRROCNo ratings yet
- Poli TraumaDocument20 pagesPoli TraumaBenjamin MalkicNo ratings yet
- Benigni Tumori Kostiju VilicaDocument32 pagesBenigni Tumori Kostiju VilicaHelena Tomas100% (1)
- Bolesti - Intrakranijalni TumoriDocument29 pagesBolesti - Intrakranijalni Tumorizixdiddy100% (1)
- Centralna SinapsaDocument16 pagesCentralna SinapsabetmenNo ratings yet
- Fiziologija SrcaDocument12 pagesFiziologija SrcaNevena BrankovicNo ratings yet
- PREZENTACIJEDocument6 pagesPREZENTACIJENickNo ratings yet
- Hromozomske AberacijeDocument1 pageHromozomske AberacijeAleksandra PericicNo ratings yet
- Auskultacija SrcaDocument8 pagesAuskultacija SrcaKemko KemsNo ratings yet
- Patologija 5Document95 pagesPatologija 5Jelena Dajana Bandovic PanicNo ratings yet
- Difuzija, Osmoza, OsmometriDocument7 pagesDifuzija, Osmoza, OsmometriMateam55No ratings yet
- Disk-Difuziona Metoda Ispitivanja Osjetljivosti Bakterija Na AntibiotikeDocument11 pagesDisk-Difuziona Metoda Ispitivanja Osjetljivosti Bakterija Na AntibiotikeMustafa Korjenić100% (3)
- Laboratorijske ProcedureDocument24 pagesLaboratorijske ProcedureJasmina Farmasi SamardžićNo ratings yet
- Samir I AnaDocument11 pagesSamir I AnaSamir CehicNo ratings yet
- Rak DojkeDocument8 pagesRak DojkeAhmet SalihovicNo ratings yet
- Koagulacija - Predavanje 2014Document18 pagesKoagulacija - Predavanje 2014vanja24312No ratings yet
- 002 Kranijalni Zivci I Putevi CnsDocument39 pages002 Kranijalni Zivci I Putevi CnsIvana Lukovic50% (2)
- Vezivno TkivoDocument17 pagesVezivno TkivoRadna Skela1No ratings yet
- Voda I Minerali Specijalizanti1Document192 pagesVoda I Minerali Specijalizanti1Dejan Todorovic100% (1)
- 1 - Prijenos Tvari Kroz Staničnu Membranu - HoDocument8 pages1 - Prijenos Tvari Kroz Staničnu Membranu - HoTamara Bluebell ﭢNo ratings yet
- FIA-Uputstvo Za Popunjavanje Uplatnice PDFDocument2 pagesFIA-Uputstvo Za Popunjavanje Uplatnice PDFabaz_zbNo ratings yet
- Bilje ĆelijeDocument4 pagesBilje ĆelijeSelma SakićNo ratings yet
- VEZBA 4 Laboratorijska DG Gljivicnih InfekcijaDocument45 pagesVEZBA 4 Laboratorijska DG Gljivicnih InfekcijaMDNo ratings yet
- Medicinski Znacaj Buke I VibracijaDocument7 pagesMedicinski Znacaj Buke I VibracijaAleksandar Aca DvornickiNo ratings yet
- Kranijalni ZivciDocument20 pagesKranijalni Zivcikarlaj97100% (1)
- Limfociti BDocument6 pagesLimfociti BIva PotočnjakNo ratings yet
- Skripta Imunogenetika Sa Osnovama ImunologijeDocument59 pagesSkripta Imunogenetika Sa Osnovama ImunologijeZerina CamdzicNo ratings yet
- Imunologija PredavanjaDocument268 pagesImunologija PredavanjascepanNo ratings yet
- Imuni Sistem, Ćelije Imunog SistemaDocument30 pagesImuni Sistem, Ćelije Imunog SistemaHany Tasha DexterNo ratings yet
- Citologija I GenetikaDocument49 pagesCitologija I GenetikaMsPaparazzyNo ratings yet