
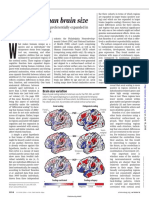
Similar Patterns of Cortical Expansion During Human Development and Evolution
Similar Patterns of Cortical Expansion During Human Development and Evolution
You might also like
- VanEssen2018-Scaling of Human Brain SizeDocument3 pagesVanEssen2018-Scaling of Human Brain Sizeiulia andreeaNo ratings yet
- Shaw Et AlDocument7 pagesShaw Et Albontes67No ratings yet
- ns2201008819 PDFDocument11 pagesns2201008819 PDFGokushimakNo ratings yet
- Near DentalDocument5 pagesNear DentalAna María Villamil CamachoNo ratings yet
- Cortical Folding PretermDocument17 pagesCortical Folding PretermmarielaNo ratings yet
- Craniofacial Growth, Maturation, and Change: Teens To MidadulthoodDocument4 pagesCraniofacial Growth, Maturation, and Change: Teens To MidadulthoodOmar DSNo ratings yet
- Attention-Deficit Hyperactivity Disorder Is Characterized by A Delay in Cortical MaturationDocument6 pagesAttention-Deficit Hyperactivity Disorder Is Characterized by A Delay in Cortical MaturationlimaNo ratings yet
- 12 Child Neurol 2015 Urger 9 20Document12 pages12 Child Neurol 2015 Urger 9 20Minerva StanciuNo ratings yet
- 2013desenvolvimento Da Microestrutura Cortical em Prematuos Chris-AnnotatedDocument6 pages2013desenvolvimento Da Microestrutura Cortical em Prematuos Chris-AnnotatedDamara GardelNo ratings yet
- PNAS 2014 Ingalhalikar 823 8Document6 pagesPNAS 2014 Ingalhalikar 823 8Akashdip SinghNo ratings yet
- 6 A Human Craniofacial Life-Course Cross-Sectionalmorphological Covariations During Postnatal Growth, Adolescence, and AgingDocument19 pages6 A Human Craniofacial Life-Course Cross-Sectionalmorphological Covariations During Postnatal Growth, Adolescence, and AgingSebastian GuevaraNo ratings yet
- PNAS2011 Petanjek NEOTsynaptsDocument6 pagesPNAS2011 Petanjek NEOTsynaptsAlejandraNo ratings yet
- The Evolution of Modern Human Brain ShapeDocument9 pagesThe Evolution of Modern Human Brain Shapelola.carvallhoNo ratings yet
- PNAS 2013 Ingalhalikar 1316909110Document6 pagesPNAS 2013 Ingalhalikar 1316909110bsaffdaberg3125100% (1)
- Neuroscience Brain AsignmentDocument23 pagesNeuroscience Brain AsignmentSyed Sajjad GillaniNo ratings yet
- Mapping Brain Maturation: Arthur W. Toga Paul M. Thompson Elizabeth R. SowellDocument13 pagesMapping Brain Maturation: Arthur W. Toga Paul M. Thompson Elizabeth R. SowellPrisiliya Van BovenNo ratings yet
- Zhou 2015Document8 pagesZhou 2015Makanudo.No ratings yet
- White Matter Development and Early Cognition in Babies and ToddlersDocument13 pagesWhite Matter Development and Early Cognition in Babies and ToddlersMariaNo ratings yet
- Jitorres LecturaOpcional Someletal.2013NatRevNeuroDocument16 pagesJitorres LecturaOpcional Someletal.2013NatRevNeuroDolosa Decencia de DecadenciaNo ratings yet
- Brainsci 13 00647Document16 pagesBrainsci 13 00647mariafernanda portoNo ratings yet
- The Evolution of Modern Human Brain Shape: ResearcharticleDocument9 pagesThe Evolution of Modern Human Brain Shape: ResearcharticleFranklinRodriguesCarlosNo ratings yet
- Van Den Bergh 2011 Li 2003: Anatomical MaturationDocument12 pagesVan Den Bergh 2011 Li 2003: Anatomical MaturationTitin Tria UtamiNo ratings yet
- Adele Diamond Close Interrelation of Motor Development and Cognitive Development Cerebellum and Prefrontal CortexDocument14 pagesAdele Diamond Close Interrelation of Motor Development and Cognitive Development Cerebellum and Prefrontal CortexAna ChicasNo ratings yet
- Physical Exercise AND NeurogenesisDocument4 pagesPhysical Exercise AND Neurogenesisesem.venezuelaNo ratings yet
- Kennedy LanguageBrainDocument17 pagesKennedy LanguageBrainPffflyers KurnawanNo ratings yet
- Frontal Lobe and Cognitive Development: Joaqu Inm - FusterDocument13 pagesFrontal Lobe and Cognitive Development: Joaqu Inm - FusterVictor BoteroNo ratings yet
- Andronikou 2014Document15 pagesAndronikou 2014JEAN CARLOS GALLO VALVERDENo ratings yet
- A Quantitative Morphometric ComparativeDocument30 pagesA Quantitative Morphometric Comparativecarmooh14No ratings yet
- Shaw Et Al. 2008Document9 pagesShaw Et Al. 2008Rodolfo Esteban Muñoz GómezNo ratings yet
- Blackwell Publishing Society For Research in Child DevelopmentDocument20 pagesBlackwell Publishing Society For Research in Child DevelopmentMatheus CorseteNo ratings yet
- 1000 DiasDocument16 pages1000 DiasNéstorNo ratings yet
- Nature - Neuro2001 - Genetics With Cover Page v2Document7 pagesNature - Neuro2001 - Genetics With Cover Page v2raghadNo ratings yet
- Differences in Frontal Network Anatomy Across Primate SpeciesDocument14 pagesDifferences in Frontal Network Anatomy Across Primate SpeciesIkhtiarNo ratings yet
- Brain Developmental Trajectories Associated With CH - 2023 - Developmental CogniDocument15 pagesBrain Developmental Trajectories Associated With CH - 2023 - Developmental CogniJuan SpinozaNo ratings yet
- What Is A Gifted Brain?: Understanding Intelligence Have We Found The Genes For Intelligence?Document35 pagesWhat Is A Gifted Brain?: Understanding Intelligence Have We Found The Genes For Intelligence?AndreiNo ratings yet
- 2017 各種sulcus測量標準化Document9 pages2017 各種sulcus測量標準化黃靖貽No ratings yet
- Coovariacion de Base y Boveda PDFDocument8 pagesCoovariacion de Base y Boveda PDFBryan Luis CARRASCAL VILLALBANo ratings yet
- Adolescent Brain Development: Iroise DumontheilDocument6 pagesAdolescent Brain Development: Iroise DumontheilJulio JaimeNo ratings yet
- Aging and The Neural Correlates of Successful Picture Encoding: Frontal Activations Compensate For Decreased Medial-Temporal ActivityDocument14 pagesAging and The Neural Correlates of Successful Picture Encoding: Frontal Activations Compensate For Decreased Medial-Temporal ActivityLau Nava-GómezNo ratings yet
- PDF MidgetDocument22 pagesPDF MidgetbagbigaloNo ratings yet
- Benign Enlargement of Extra-Axial SpaceDocument7 pagesBenign Enlargement of Extra-Axial SpaceAlfredo CaceresNo ratings yet
- Fped 05 00073Document9 pagesFped 05 00073Silvina DurhandNo ratings yet
- Joa 235 432Document20 pagesJoa 235 432ElCanaldeSaadNo ratings yet
- 1 s2.0 B9780123747488000068 MainDocument25 pages1 s2.0 B9780123747488000068 MainAlex SamuelNo ratings yet
- Chapter 15 - Development of The Zebrafish Pronep - 2020 - The Zebrafish in BiomeDocument6 pagesChapter 15 - Development of The Zebrafish Pronep - 2020 - The Zebrafish in BiomeNicolas BaronNo ratings yet
- Limperopoulos Bolduc09 Neurodevelopmental Outcomes After CB Injury REVIEWDocument12 pagesLimperopoulos Bolduc09 Neurodevelopmental Outcomes After CB Injury REVIEWEccoNo ratings yet
- Neuroanatomic Alterations and Social and Communication Deficits in Monozygotic Twins Discordant For Autism DisorderDocument9 pagesNeuroanatomic Alterations and Social and Communication Deficits in Monozygotic Twins Discordant For Autism DisorderChristianLambertNo ratings yet
- Article 3Document11 pagesArticle 3moodyxNo ratings yet
- Sex Differences 1Document6 pagesSex Differences 1RocioMorenoTarazonaNo ratings yet
- Brain Volume and Shape in Infants With Deformational PlagiocephalyDocument8 pagesBrain Volume and Shape in Infants With Deformational PlagiocephalychiaraNo ratings yet
- Gene Expression Analysis Reveals The Tipping Points During Infant Brain Development For Human and ChimpanzeeDocument10 pagesGene Expression Analysis Reveals The Tipping Points During Infant Brain Development For Human and ChimpanzeeCarlos IsacazNo ratings yet
- A Morphometric Comparison of The Parietal Lobe in Modern Humans and Neanderthals 2020 32c 628hDocument11 pagesA Morphometric Comparison of The Parietal Lobe in Modern Humans and Neanderthals 2020 32c 628hlola.carvallhoNo ratings yet
- Cortical Surface Area of The Left Frontal Pole Is Associated With Visuospatial Working Memory CapacityDocument6 pagesCortical Surface Area of The Left Frontal Pole Is Associated With Visuospatial Working Memory CapacityManal ChennoufNo ratings yet
- Perspectives On The Use of The Baboon in Embryology and Teratology ResearchDocument18 pagesPerspectives On The Use of The Baboon in Embryology and Teratology ResearchYusraNo ratings yet
- Human Brain Biochemistry: American Journal of BioscienceDocument13 pagesHuman Brain Biochemistry: American Journal of BioscienceEugen MNo ratings yet
- Sex Differences in Spatial Navigation and Perception in Human Adolescents and Emerging AdultsDocument9 pagesSex Differences in Spatial Navigation and Perception in Human Adolescents and Emerging AdultsMusliminNo ratings yet
- Environmental Influences On The Pace of Brain DevelopmentDocument13 pagesEnvironmental Influences On The Pace of Brain DevelopmentShekel SibalaNo ratings yet
- Bruner 2021Document19 pagesBruner 2021Manuel CuminiNo ratings yet


























































Download as pdf or txt
You might also like
- VanEssen2018-Scaling of Human Brain SizeDocument3 pagesVanEssen2018-Scaling of Human Brain Sizeiulia andreeaNo ratings yet
- Shaw Et AlDocument7 pagesShaw Et Albontes67No ratings yet
- ns2201008819 PDFDocument11 pagesns2201008819 PDFGokushimakNo ratings yet
- Near DentalDocument5 pagesNear DentalAna María Villamil CamachoNo ratings yet
- Cortical Folding PretermDocument17 pagesCortical Folding PretermmarielaNo ratings yet
- Craniofacial Growth, Maturation, and Change: Teens To MidadulthoodDocument4 pagesCraniofacial Growth, Maturation, and Change: Teens To MidadulthoodOmar DSNo ratings yet
- Attention-Deficit Hyperactivity Disorder Is Characterized by A Delay in Cortical MaturationDocument6 pagesAttention-Deficit Hyperactivity Disorder Is Characterized by A Delay in Cortical MaturationlimaNo ratings yet
- 12 Child Neurol 2015 Urger 9 20Document12 pages12 Child Neurol 2015 Urger 9 20Minerva StanciuNo ratings yet
- 2013desenvolvimento Da Microestrutura Cortical em Prematuos Chris-AnnotatedDocument6 pages2013desenvolvimento Da Microestrutura Cortical em Prematuos Chris-AnnotatedDamara GardelNo ratings yet
- PNAS 2014 Ingalhalikar 823 8Document6 pagesPNAS 2014 Ingalhalikar 823 8Akashdip SinghNo ratings yet
- 6 A Human Craniofacial Life-Course Cross-Sectionalmorphological Covariations During Postnatal Growth, Adolescence, and AgingDocument19 pages6 A Human Craniofacial Life-Course Cross-Sectionalmorphological Covariations During Postnatal Growth, Adolescence, and AgingSebastian GuevaraNo ratings yet
- PNAS2011 Petanjek NEOTsynaptsDocument6 pagesPNAS2011 Petanjek NEOTsynaptsAlejandraNo ratings yet
- The Evolution of Modern Human Brain ShapeDocument9 pagesThe Evolution of Modern Human Brain Shapelola.carvallhoNo ratings yet
- PNAS 2013 Ingalhalikar 1316909110Document6 pagesPNAS 2013 Ingalhalikar 1316909110bsaffdaberg3125100% (1)
- Neuroscience Brain AsignmentDocument23 pagesNeuroscience Brain AsignmentSyed Sajjad GillaniNo ratings yet
- Mapping Brain Maturation: Arthur W. Toga Paul M. Thompson Elizabeth R. SowellDocument13 pagesMapping Brain Maturation: Arthur W. Toga Paul M. Thompson Elizabeth R. SowellPrisiliya Van BovenNo ratings yet
- Zhou 2015Document8 pagesZhou 2015Makanudo.No ratings yet
- White Matter Development and Early Cognition in Babies and ToddlersDocument13 pagesWhite Matter Development and Early Cognition in Babies and ToddlersMariaNo ratings yet
- Jitorres LecturaOpcional Someletal.2013NatRevNeuroDocument16 pagesJitorres LecturaOpcional Someletal.2013NatRevNeuroDolosa Decencia de DecadenciaNo ratings yet
- Brainsci 13 00647Document16 pagesBrainsci 13 00647mariafernanda portoNo ratings yet
- The Evolution of Modern Human Brain Shape: ResearcharticleDocument9 pagesThe Evolution of Modern Human Brain Shape: ResearcharticleFranklinRodriguesCarlosNo ratings yet
- Van Den Bergh 2011 Li 2003: Anatomical MaturationDocument12 pagesVan Den Bergh 2011 Li 2003: Anatomical MaturationTitin Tria UtamiNo ratings yet
- Adele Diamond Close Interrelation of Motor Development and Cognitive Development Cerebellum and Prefrontal CortexDocument14 pagesAdele Diamond Close Interrelation of Motor Development and Cognitive Development Cerebellum and Prefrontal CortexAna ChicasNo ratings yet
- Physical Exercise AND NeurogenesisDocument4 pagesPhysical Exercise AND Neurogenesisesem.venezuelaNo ratings yet
- Kennedy LanguageBrainDocument17 pagesKennedy LanguageBrainPffflyers KurnawanNo ratings yet
- Frontal Lobe and Cognitive Development: Joaqu Inm - FusterDocument13 pagesFrontal Lobe and Cognitive Development: Joaqu Inm - FusterVictor BoteroNo ratings yet
- Andronikou 2014Document15 pagesAndronikou 2014JEAN CARLOS GALLO VALVERDENo ratings yet
- A Quantitative Morphometric ComparativeDocument30 pagesA Quantitative Morphometric Comparativecarmooh14No ratings yet
- Shaw Et Al. 2008Document9 pagesShaw Et Al. 2008Rodolfo Esteban Muñoz GómezNo ratings yet
- Blackwell Publishing Society For Research in Child DevelopmentDocument20 pagesBlackwell Publishing Society For Research in Child DevelopmentMatheus CorseteNo ratings yet
- 1000 DiasDocument16 pages1000 DiasNéstorNo ratings yet
- Nature - Neuro2001 - Genetics With Cover Page v2Document7 pagesNature - Neuro2001 - Genetics With Cover Page v2raghadNo ratings yet
- Differences in Frontal Network Anatomy Across Primate SpeciesDocument14 pagesDifferences in Frontal Network Anatomy Across Primate SpeciesIkhtiarNo ratings yet
- Brain Developmental Trajectories Associated With CH - 2023 - Developmental CogniDocument15 pagesBrain Developmental Trajectories Associated With CH - 2023 - Developmental CogniJuan SpinozaNo ratings yet
- What Is A Gifted Brain?: Understanding Intelligence Have We Found The Genes For Intelligence?Document35 pagesWhat Is A Gifted Brain?: Understanding Intelligence Have We Found The Genes For Intelligence?AndreiNo ratings yet
- 2017 各種sulcus測量標準化Document9 pages2017 各種sulcus測量標準化黃靖貽No ratings yet
- Coovariacion de Base y Boveda PDFDocument8 pagesCoovariacion de Base y Boveda PDFBryan Luis CARRASCAL VILLALBANo ratings yet
- Adolescent Brain Development: Iroise DumontheilDocument6 pagesAdolescent Brain Development: Iroise DumontheilJulio JaimeNo ratings yet
- Aging and The Neural Correlates of Successful Picture Encoding: Frontal Activations Compensate For Decreased Medial-Temporal ActivityDocument14 pagesAging and The Neural Correlates of Successful Picture Encoding: Frontal Activations Compensate For Decreased Medial-Temporal ActivityLau Nava-GómezNo ratings yet
- PDF MidgetDocument22 pagesPDF MidgetbagbigaloNo ratings yet
- Benign Enlargement of Extra-Axial SpaceDocument7 pagesBenign Enlargement of Extra-Axial SpaceAlfredo CaceresNo ratings yet
- Fped 05 00073Document9 pagesFped 05 00073Silvina DurhandNo ratings yet
- Joa 235 432Document20 pagesJoa 235 432ElCanaldeSaadNo ratings yet
- 1 s2.0 B9780123747488000068 MainDocument25 pages1 s2.0 B9780123747488000068 MainAlex SamuelNo ratings yet
- Chapter 15 - Development of The Zebrafish Pronep - 2020 - The Zebrafish in BiomeDocument6 pagesChapter 15 - Development of The Zebrafish Pronep - 2020 - The Zebrafish in BiomeNicolas BaronNo ratings yet
- Limperopoulos Bolduc09 Neurodevelopmental Outcomes After CB Injury REVIEWDocument12 pagesLimperopoulos Bolduc09 Neurodevelopmental Outcomes After CB Injury REVIEWEccoNo ratings yet
- Neuroanatomic Alterations and Social and Communication Deficits in Monozygotic Twins Discordant For Autism DisorderDocument9 pagesNeuroanatomic Alterations and Social and Communication Deficits in Monozygotic Twins Discordant For Autism DisorderChristianLambertNo ratings yet
- Article 3Document11 pagesArticle 3moodyxNo ratings yet
- Sex Differences 1Document6 pagesSex Differences 1RocioMorenoTarazonaNo ratings yet
- Brain Volume and Shape in Infants With Deformational PlagiocephalyDocument8 pagesBrain Volume and Shape in Infants With Deformational PlagiocephalychiaraNo ratings yet
- Gene Expression Analysis Reveals The Tipping Points During Infant Brain Development For Human and ChimpanzeeDocument10 pagesGene Expression Analysis Reveals The Tipping Points During Infant Brain Development For Human and ChimpanzeeCarlos IsacazNo ratings yet
- A Morphometric Comparison of The Parietal Lobe in Modern Humans and Neanderthals 2020 32c 628hDocument11 pagesA Morphometric Comparison of The Parietal Lobe in Modern Humans and Neanderthals 2020 32c 628hlola.carvallhoNo ratings yet
- Cortical Surface Area of The Left Frontal Pole Is Associated With Visuospatial Working Memory CapacityDocument6 pagesCortical Surface Area of The Left Frontal Pole Is Associated With Visuospatial Working Memory CapacityManal ChennoufNo ratings yet
- Perspectives On The Use of The Baboon in Embryology and Teratology ResearchDocument18 pagesPerspectives On The Use of The Baboon in Embryology and Teratology ResearchYusraNo ratings yet
- Human Brain Biochemistry: American Journal of BioscienceDocument13 pagesHuman Brain Biochemistry: American Journal of BioscienceEugen MNo ratings yet
- Sex Differences in Spatial Navigation and Perception in Human Adolescents and Emerging AdultsDocument9 pagesSex Differences in Spatial Navigation and Perception in Human Adolescents and Emerging AdultsMusliminNo ratings yet
- Environmental Influences On The Pace of Brain DevelopmentDocument13 pagesEnvironmental Influences On The Pace of Brain DevelopmentShekel SibalaNo ratings yet
- Bruner 2021Document19 pagesBruner 2021Manuel CuminiNo ratings yet