Download as pdf or txt
You might also like
- Experiments in Molecular Cell Biology: A Problems Book With Multiple-Choice Question-Based TestsDocument20 pagesExperiments in Molecular Cell Biology: A Problems Book With Multiple-Choice Question-Based TestsKudumarNo ratings yet
- Forensics Lab ReportDocument15 pagesForensics Lab Reportachita450No ratings yet
- Post Translational ModificationsDocument35 pagesPost Translational ModificationsGandhiraj VijayaragavanNo ratings yet
- Lecture 3 - Chapter 8-Cytoskeleton ADocument75 pagesLecture 3 - Chapter 8-Cytoskeleton AKw Chan33% (3)
- Protein FunctionDocument39 pagesProtein FunctionDeana Namirembe100% (1)
- Biochemical Energetics: Peranan Atp BioenergetikaDocument23 pagesBiochemical Energetics: Peranan Atp BioenergetikaHerryNo ratings yet
- Konsep Dasar Genetika KedokteranDocument40 pagesKonsep Dasar Genetika KedokteranSarah Nadia RasidiNo ratings yet
- Text Book of EmbryologyDocument476 pagesText Book of EmbryologyJojin Jose100% (2)
- CytoskeletonDocument86 pagesCytoskeletonAbid Al RezaNo ratings yet
- lectut-BTN-303-pdf-Protoplast Technology PDFDocument12 pageslectut-BTN-303-pdf-Protoplast Technology PDFqwertNo ratings yet
- Euchromatin and Heterochromatin PDFDocument2 pagesEuchromatin and Heterochromatin PDFRachelNo ratings yet
- Gene ExpressionDocument23 pagesGene ExpressionSutama ArtaNo ratings yet
- Fish Reproduction: 1 AnatomyDocument11 pagesFish Reproduction: 1 Anatomysksingl350No ratings yet
- anatomy 8 scalp تعلم بذكاءDocument8 pagesanatomy 8 scalp تعلم بذكاءmahmoud ezzeldeenNo ratings yet
- Limb DevelopmentDocument95 pagesLimb Developmentcandy_kb17100% (1)
- Metabolic OR Endocrine Disorders: Amanda Du, MD, DPPS, DPSNBM Pediatrician-NeonatologistDocument124 pagesMetabolic OR Endocrine Disorders: Amanda Du, MD, DPPS, DPSNBM Pediatrician-NeonatologistLyn LynNo ratings yet
- Protein Synthesis (Transcription and Translation) : Dr. Marhaen Hardjo, M.Biomed, PHDDocument159 pagesProtein Synthesis (Transcription and Translation) : Dr. Marhaen Hardjo, M.Biomed, PHDsNo ratings yet
- 1 Red Blood Cells Anemia and PolycythemiaDocument43 pages1 Red Blood Cells Anemia and PolycythemiaGeevee Naganag VentulaNo ratings yet
- Wiley - Gene Cloning and DNA Analysis An Introduction, 7th Edition - 978-1-119-07256-0Document2 pagesWiley - Gene Cloning and DNA Analysis An Introduction, 7th Edition - 978-1-119-07256-0alfiNo ratings yet
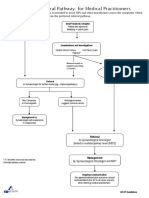
- Cervical Cancer Referral PathwayDocument1 pageCervical Cancer Referral PathwaySifa AditNo ratings yet
- Teknik Analisis Biologi Moleku PDFDocument40 pagesTeknik Analisis Biologi Moleku PDFACHMAD HAIRIL AffanNo ratings yet
- Genetics of Cystic FibrosisDocument3 pagesGenetics of Cystic Fibrosislutfiah_liaNo ratings yet
- Nomina Anatomica Veterinaria 2012 PDFDocument177 pagesNomina Anatomica Veterinaria 2012 PDFFernando CruzNo ratings yet
- Apoptosis ManualDocument174 pagesApoptosis Manualbibeksd796No ratings yet
- PCR An OverviewDocument41 pagesPCR An OverviewnassradabourNo ratings yet
- Embryology - Updates and Highlights On Classic TopicsDocument222 pagesEmbryology - Updates and Highlights On Classic TopicsIndera Vyas100% (1)
- Protein StructureDocument41 pagesProtein Structureteklay100% (1)
- Veterinary Basics1Document119 pagesVeterinary Basics1Saleem Ahmed Shahwani0% (1)
- 16) Signaling Pathways That Control Gene ActivityDocument27 pages16) Signaling Pathways That Control Gene ActivityRNo ratings yet
- 14.kuliah-Histologi MataDocument35 pages14.kuliah-Histologi MataRahmat NugrohoNo ratings yet
- Oogenesis and The Ovarian Cycle: Angelbert R. Bacongco Justin Rhea M. BandiolaDocument19 pagesOogenesis and The Ovarian Cycle: Angelbert R. Bacongco Justin Rhea M. BandiolaJustin Rhea Bandiola100% (1)
- Omics Technology: October 2010Document28 pagesOmics Technology: October 2010Janescu LucianNo ratings yet
- Polycistronic and Monocistronic mRNAsDocument7 pagesPolycistronic and Monocistronic mRNAskailas ambadi100% (1)
- Introduction To HistologyDocument32 pagesIntroduction To HistologyPushparaj ShettyNo ratings yet
- Alkaline PhosphataseDocument19 pagesAlkaline PhosphataseBernardo JoshuaNo ratings yet
- SOAL Masuk 2018Document142 pagesSOAL Masuk 2018Yoza FirdaozNo ratings yet
- ABDocument22 pagesABpk 77No ratings yet
- Receptor Tyrosine Kinase InhibitorsDocument63 pagesReceptor Tyrosine Kinase InhibitorsNoor Nour100% (1)
- Regional Anesthesia - 3B NotesDocument5 pagesRegional Anesthesia - 3B NotesBetina HernandezNo ratings yet
- Seven Jump Dalam Diskusi TutorialDocument39 pagesSeven Jump Dalam Diskusi TutorialkadinfathiaNo ratings yet
- Biology Lecture, Chapter 5Document84 pagesBiology Lecture, Chapter 5Nick GoldingNo ratings yet
- Biochemistry of Bone & Muscle: Dr. Syahrijuita, M.Kes, SP - THT-KLDocument69 pagesBiochemistry of Bone & Muscle: Dr. Syahrijuita, M.Kes, SP - THT-KLyantiNo ratings yet
- Discuss The Role of Immunology in SurgeryDocument16 pagesDiscuss The Role of Immunology in SurgeryrosybashNo ratings yet
- Cell Biology LabManual Version 8.0 May 2012 1 TRUNCATEDDocument57 pagesCell Biology LabManual Version 8.0 May 2012 1 TRUNCATEDதுர்காஸ்ரீ கங்கா ராதிகாNo ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
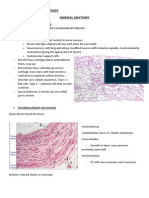
- Veterinary Normal HistologyDocument10 pagesVeterinary Normal HistologyIshtar Katherine ChanNo ratings yet
- Histology of Lymphoid Organ PDFDocument82 pagesHistology of Lymphoid Organ PDFRionaldy TaminNo ratings yet
- Brain Normal PDFDocument64 pagesBrain Normal PDFRicky Chupruut ArisandyNo ratings yet
- Factsheet - Livestock - ReproductiveDiseases in Cattle v2Document3 pagesFactsheet - Livestock - ReproductiveDiseases in Cattle v2lenn chandNo ratings yet
- Third Week of DevelopmentDocument34 pagesThird Week of DevelopmentAriba Asif100% (1)
- Pedi Pearls 2 - Scrubed PDFDocument46 pagesPedi Pearls 2 - Scrubed PDFthomasfx10100% (1)
- Surgery of The Bovine Large IntestineDocument18 pagesSurgery of The Bovine Large IntestineJhon Bustamante CanoNo ratings yet
- Congenital and Genetic DiseasesDocument84 pagesCongenital and Genetic Diseasesdrlandge100% (1)
- Genetics DisorderDocument42 pagesGenetics DisorderRoshan jaiswalNo ratings yet
- Post Transcriptional ModificationDocument20 pagesPost Transcriptional ModificationZain YaqoobNo ratings yet
- Molecular BiotechnologyDocument40 pagesMolecular BiotechnologyBhaskar GangulyNo ratings yet
- Usg 20088Document18 pagesUsg 20088iridaNo ratings yet
- Fetal Biophysical Profile. A Critical Appraisal PDFDocument11 pagesFetal Biophysical Profile. A Critical Appraisal PDFchronos6534No ratings yet
- Principles of Stem Cell Biology and Cancer: Future Applications and TherapeuticsFrom EverandPrinciples of Stem Cell Biology and Cancer: Future Applications and TherapeuticsNo ratings yet
- Chapter 7Document40 pagesChapter 7NurulJannah ARNo ratings yet
- Transposons Mediated MutagenesisDocument41 pagesTransposons Mediated Mutagenesissaurabh784No ratings yet
- Genome Bioinformatics: Richard Durbin Sanger Institute Rd@sanger - Ac.ukDocument49 pagesGenome Bioinformatics: Richard Durbin Sanger Institute Rd@sanger - Ac.ukAshishNo ratings yet
- Study Guide For CHP 15Document4 pagesStudy Guide For CHP 15Fati UmaruNo ratings yet
- Transposable Elements & Transposable Elements in Bacteria: Insertion Sequences (IS)Document22 pagesTransposable Elements & Transposable Elements in Bacteria: Insertion Sequences (IS)Gayathri deviNo ratings yet
- BT2030 NolDocument51 pagesBT2030 Nolkrishradhan1100% (1)
- Genetic RecombinationDocument55 pagesGenetic RecombinationRiyaniMartyasariNo ratings yet
- Sanmiguel, 1998Document8 pagesSanmiguel, 1998Chie SakuraNo ratings yet
- bt301 Full Book McqsDocument31 pagesbt301 Full Book McqsAarish AliNo ratings yet
- 2019MackinSJMPhD PDFDocument184 pages2019MackinSJMPhD PDFLuciani Magdalena Palacios GarcíaNo ratings yet
- Molecular Plant Taxonomy. Methods in Molecular Biology (2021)Document407 pagesMolecular Plant Taxonomy. Methods in Molecular Biology (2021)Giovanni Celso Paton MamaniNo ratings yet
- Classif lncRNADocument13 pagesClassif lncRNAoli.finetNo ratings yet
- (Learning Materials in Biosciences) Melanie Kappelmann-Fenzl - Next Generation Sequencing and Data Analysis-Springer (2021)Document225 pages(Learning Materials in Biosciences) Melanie Kappelmann-Fenzl - Next Generation Sequencing and Data Analysis-Springer (2021)gndfbj100% (2)
- Transposable Elements, EpigeneticsDocument10 pagesTransposable Elements, EpigeneticsAlejandro López OchoaNo ratings yet
- Microsatellites: Simple Sequences With Complex Evolution: Hans EllegrenDocument11 pagesMicrosatellites: Simple Sequences With Complex Evolution: Hans Ellegrenkrista_jw9525No ratings yet
- Mapping and Analysis of The LINE and SINE Type of Repetitive Elements in RiceDocument4 pagesMapping and Analysis of The LINE and SINE Type of Repetitive Elements in RiceBrijesh Singh YadavNo ratings yet
- Aspergillus Fumigatus and AspergillosisDocument41 pagesAspergillus Fumigatus and AspergillosisNurul FadilahNo ratings yet
- Intelligence N GenesDocument208 pagesIntelligence N Genestoussman80No ratings yet
- Lecture 1 - Genes and GenomicsDocument51 pagesLecture 1 - Genes and GenomicsDeepali SinghNo ratings yet
- El Sayed2005Document7 pagesEl Sayed2005Romeo Pomarí JuárezNo ratings yet
- Feschotte 2007Document38 pagesFeschotte 2007Axel GómezNo ratings yet
- Natural History of Eukaryotic DNA Methylation SystemsDocument80 pagesNatural History of Eukaryotic DNA Methylation SystemsDumbo MkuNo ratings yet
- Human Genome ProjectDocument33 pagesHuman Genome ProjectFatima IffatNo ratings yet
- Transposons Mobile DNADocument6 pagesTransposons Mobile DNAkerkour-abd1523No ratings yet
- Basic Cell and Molecular Biology 4e - What We Know and How Found ODocument629 pagesBasic Cell and Molecular Biology 4e - What We Know and How Found Oмакс100% (1)
- MutagenDocument15 pagesMutagenDibyakNo ratings yet
- Chinhoyi University of Technology: Biotechnology DepartmentDocument8 pagesChinhoyi University of Technology: Biotechnology DepartmentDumisani Nguni100% (1)
- Pv92 Student InstructionsDocument20 pagesPv92 Student Instructionsjugdeep15No ratings yet
- Reverse TranscriptionDocument9 pagesReverse TranscriptionzahraNo ratings yet