
Algal Flagella PDF
Algal Flagella PDF
You might also like
- Cell Modification ModuleDocument30 pagesCell Modification ModuleDuane100% (1)
- USMLE World: Revision QuestionsDocument65 pagesUSMLE World: Revision QuestionsDustin91% (23)
- Phylum Protozoa 02Document25 pagesPhylum Protozoa 02Haris AyubNo ratings yet
- Cilia and FlagellaDocument25 pagesCilia and FlagellaLia Savitri RomdaniNo ratings yet
- Cilia and Flagella: Structure and Function in EukaryotesDocument10 pagesCilia and Flagella: Structure and Function in EukaryotesEr Purushottam PalNo ratings yet
- Science Wps OfficeDocument16 pagesScience Wps OfficeLuke Chase JacintoNo ratings yet
- Locomotion in EuglenaDocument4 pagesLocomotion in Euglenamanojitchatterjee2007No ratings yet
- Locomotary Organelles in ProtozoaDocument25 pagesLocomotary Organelles in Protozoaashley clintonNo ratings yet
- Structure of Cilia and Flagella: - by Neha LoharDocument18 pagesStructure of Cilia and Flagella: - by Neha Loharjoy parimalaNo ratings yet
- Cilia and FlagellaDocument8 pagesCilia and Flagellaamnamalik1304No ratings yet
- Ultrastructure and Development of The Flagellar Apparatus and Flagellar Motion in The Colonial Green Alga AstrephomeneDocument22 pagesUltrastructure and Development of The Flagellar Apparatus and Flagellar Motion in The Colonial Green Alga AstrephomeneARMY SistersNo ratings yet
- Cell Membrane Unit 1-2Document52 pagesCell Membrane Unit 1-2Kemoy FrancisNo ratings yet
- Nihms 367187Document14 pagesNihms 367187Tiego FerreiraNo ratings yet
- Invertebrate Anatomy LengkapDocument49 pagesInvertebrate Anatomy LengkapMayola ArdaNo ratings yet
- Cilia and FlagellaDocument4 pagesCilia and FlagellaAnurag chavan100% (1)
- FlagellumDocument13 pagesFlagellumDaner BaxhijaNo ratings yet
- Structure and FunctionDocument12 pagesStructure and FunctionJustine PamaNo ratings yet
- Semester 1 Locomotion in ProtozoaDocument2 pagesSemester 1 Locomotion in Protozoaashwinirana8No ratings yet
- Microtubules: Organization: Single Microtubules Are Distributed Within The Cytoplasm. Also, MicrotubulesDocument6 pagesMicrotubules: Organization: Single Microtubules Are Distributed Within The Cytoplasm. Also, MicrotubulesArif NabeelNo ratings yet
- Flagellar Restoration in C. ReinhardiDocument8 pagesFlagellar Restoration in C. Reinhardinasek22No ratings yet
- The Cell: Prokaryotic EukaryoticDocument28 pagesThe Cell: Prokaryotic EukaryoticMastersBestHackersNo ratings yet
- The CellDocument24 pagesThe CellCaren AdongoNo ratings yet
- Full Download Zoology 9th Edition Miller Solutions ManualDocument35 pagesFull Download Zoology 9th Edition Miller Solutions Manualcaveneywilliams100% (19)
- 2 PARA 1 - Protozoa - FlagellatesDocument13 pages2 PARA 1 - Protozoa - FlagellatesTricia LlorinNo ratings yet
- Exercise 2: Cells: Examining Cyanobacteria (Blue Green Algae)Document5 pagesExercise 2: Cells: Examining Cyanobacteria (Blue Green Algae)acgonzales28No ratings yet
- Opposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumDocument8 pagesOpposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumFernandoNo ratings yet
- Biol120 Lecture 7Document55 pagesBiol120 Lecture 7mooosemnNo ratings yet
- Lab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeDocument4 pagesLab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeMichael TimsonNo ratings yet
- Morphology and Fine StructureDocument67 pagesMorphology and Fine StructureDiksha PadamNo ratings yet
- The Dynein Family at A GlanceDocument3 pagesThe Dynein Family at A GlanceNovVie VietTha Sccor IINo ratings yet
- EuglenaDocument4 pagesEuglenaBio SciencesNo ratings yet
- Biomed 05Document12 pagesBiomed 05api-3706483No ratings yet
- Module (Cell Nucleus)Document12 pagesModule (Cell Nucleus)Edgie JunelaNo ratings yet
- Parts of A Cell BIOLOGYDocument43 pagesParts of A Cell BIOLOGYSam Ferbyn LomotanNo ratings yet
- Cell and MolecularDocument37 pagesCell and MolecularAbhishek AbhiNo ratings yet
- 1-Ribosomes: Nucleolus Nuclear PoresDocument12 pages1-Ribosomes: Nucleolus Nuclear Poresapi-3706483No ratings yet
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-530699294No ratings yet
- Chapter 2 NeuronDocument49 pagesChapter 2 NeuronAfNo ratings yet
- The Cell: by DR - Hikmat Fatima HashmiDocument53 pagesThe Cell: by DR - Hikmat Fatima Hashmibeverly brittoNo ratings yet
- CellDocument41 pagesCellAditya VermaNo ratings yet
- Cell Unit of Life Question AnswerDocument8 pagesCell Unit of Life Question Answershafiahmad2014.15No ratings yet
- Ultrastructure of Bacteria & EndosporeDocument8 pagesUltrastructure of Bacteria & EndosporeSubrata KunduNo ratings yet
- Kinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardDocument11 pagesKinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardAlessandro BianchessiNo ratings yet
- Elsc Easy Learn Singh Classes: Cell OrganelleDocument13 pagesElsc Easy Learn Singh Classes: Cell OrganelleNandini SinghNo ratings yet
- CH 6 BioDocument5 pagesCH 6 BioedwarddameNo ratings yet
- Cells: H1N3 Publications SDN BHD®Document22 pagesCells: H1N3 Publications SDN BHD®Noor HafizNo ratings yet
- CellsDocument46 pagesCellsErfanShekarrizNo ratings yet
- Organelle: Flagellum DefinitionDocument4 pagesOrganelle: Flagellum DefinitionMadhanNo ratings yet
- Eukaryotic CellsDocument21 pagesEukaryotic Cellsjaysenires13tkdNo ratings yet
- Britannica Print DemoDocument9 pagesBritannica Print DemoduongtuScribdNo ratings yet
- Organelles Working TogetherDocument4 pagesOrganelles Working TogetherVilma Malaya VisarioNo ratings yet
- Assignment II: Macaranas, Mar SDocument7 pagesAssignment II: Macaranas, Mar SMar MacaranasNo ratings yet
- GROUP 5 PhysioLab Activity 3Document7 pagesGROUP 5 PhysioLab Activity 3Deasserei TatelNo ratings yet
- Biochem AssignmentDocument2 pagesBiochem AssignmentAngel GoNo ratings yet
- Lecture 4 Notes 2013Document4 pagesLecture 4 Notes 2013Richard HampsonNo ratings yet
- Cells: A. Ultrastructure & Function of The CellDocument9 pagesCells: A. Ultrastructure & Function of The CellRhea Lyn LamosteNo ratings yet
- Cytoskeleton: Jetro P. MagnayonDocument66 pagesCytoskeleton: Jetro P. MagnayonChristian Santiago100% (1)
- Lab Report 5Document9 pagesLab Report 5jvelezdanielaNo ratings yet
- Cellular LocomotionDocument14 pagesCellular LocomotionEshaNo ratings yet
- Cell Biology Practical 3 PDFDocument11 pagesCell Biology Practical 3 PDFEswaran Singam100% (1)
- Flagella and MotilityDocument17 pagesFlagella and Motilityqasim mughaNo ratings yet
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceFrom EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceNo ratings yet
- Plant Macro-And Micronutrient MineralsDocument5 pagesPlant Macro-And Micronutrient Mineralsmanoj_rkl_07No ratings yet
- Electroporation: Jac A NickoloffDocument3 pagesElectroporation: Jac A Nickoloffmanoj_rkl_07No ratings yet
- DNA Damage: Paul W DoetschDocument7 pagesDNA Damage: Paul W Doetschmanoj_rkl_07No ratings yet
- Phyllosphere PDFDocument8 pagesPhyllosphere PDFmanoj_rkl_07No ratings yet
- Dideoxy Sequencing of DNA PDFDocument16 pagesDideoxy Sequencing of DNA PDFmanoj_rkl_07No ratings yet
- Genetic Code Introduction PDFDocument10 pagesGenetic Code Introduction PDFmanoj_rkl_07No ratings yet
- Closteroviridae: Historical PerspectiveDocument6 pagesClosteroviridae: Historical Perspectivemanoj_rkl_07No ratings yet
- Root Nodules (Rhizobium Legumes) PDFDocument2 pagesRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07No ratings yet
- Terpenoids Lower PDFDocument7 pagesTerpenoids Lower PDFmanoj_rkl_07No ratings yet
- RNA Plant and Animal Virus Replication PDFDocument9 pagesRNA Plant and Animal Virus Replication PDFmanoj_rkl_07No ratings yet
- Plant Tracheary Elements PDFDocument2 pagesPlant Tracheary Elements PDFmanoj_rkl_07No ratings yet
- S Adenosylmethionine PDFDocument7 pagesS Adenosylmethionine PDFmanoj_rkl_07No ratings yet
- Plant Water Relations PDFDocument7 pagesPlant Water Relations PDFmanoj_rkl_07No ratings yet
- Plant Cytoskeleton PDFDocument7 pagesPlant Cytoskeleton PDFmanoj_rkl_07No ratings yet
- Plant Genome Projects PDFDocument4 pagesPlant Genome Projects PDFmanoj_rkl_07100% (1)
- Plant Cells Peroxisomes and Glyoxysomes PDFDocument7 pagesPlant Cells Peroxisomes and Glyoxysomes PDFmanoj_rkl_07No ratings yet
- Cell - The Unit of Life DPP 09 Yakeen NEET 2.0 2025 (LegDocument3 pagesCell - The Unit of Life DPP 09 Yakeen NEET 2.0 2025 (LegBISWARANJAN SAHOONo ratings yet
- Chest Injury and Its TypesDocument14 pagesChest Injury and Its TypesKoochi PoojithaNo ratings yet
- Cell Theory: Biology Scientific Theory CellsDocument5 pagesCell Theory: Biology Scientific Theory CellsNiel S. DefensorNo ratings yet
- Case Studies in Veterinary ImmuneologyDocument209 pagesCase Studies in Veterinary ImmuneologyNathalia gallegoNo ratings yet
- Microbial Morphology and TaxonomyDocument6 pagesMicrobial Morphology and TaxonomyJasmin Pearl AndayaNo ratings yet
- Locomotary Organelles in ProtozoaDocument25 pagesLocomotary Organelles in Protozoaashley clintonNo ratings yet
- Histology and Cell Biology An Introduction To Pathology 5Th Edition Abraham L Kierszenbaum M D PH D Full ChapterDocument67 pagesHistology and Cell Biology An Introduction To Pathology 5Th Edition Abraham L Kierszenbaum M D PH D Full Chapterdennis.roberson462100% (17)
- Cell Organelles - Biology Class 11 - NEETDocument18 pagesCell Organelles - Biology Class 11 - NEETsanjNo ratings yet
- Ws 2 Answer Key Unit 2 22-23Document9 pagesWs 2 Answer Key Unit 2 22-23majdiabd5678No ratings yet
- Test Bank For Color Textbook of Histology 3rd Edition Leslie P GartnerDocument12 pagesTest Bank For Color Textbook of Histology 3rd Edition Leslie P Gartnerbrennaquani2mNo ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- Chapter 6Document13 pagesChapter 6gahaganeNo ratings yet
- Ncert Highlight - Cell - by Seep PahujaDocument18 pagesNcert Highlight - Cell - by Seep PahujaskindustrieshelplineNo ratings yet
- Vol.14 No.3 PDFDocument140 pagesVol.14 No.3 PDFAnn JoseNo ratings yet
- BIOL 102 Lab Manual 2022 - 2023Document184 pagesBIOL 102 Lab Manual 2022 - 2023Saya SinNo ratings yet
- 03 Cell BiologyDocument111 pages03 Cell BiologyPrincess Noreen SavellanoNo ratings yet
- Fragrance Chemistry The Science of The Sense of SmellDocument631 pagesFragrance Chemistry The Science of The Sense of SmellGuedri Ghania0% (1)
- Cell Organelles DefinitionDocument40 pagesCell Organelles Definitionmuhammad aslam0% (1)
- Specialised Cells-& Levels of OrganisationsDocument34 pagesSpecialised Cells-& Levels of OrganisationsEmna MbarekNo ratings yet
- Specialised Cells Information SheetsDocument8 pagesSpecialised Cells Information SheetsVy LyNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- 3F Sta. Cruz Bldg. Cor. Fuentes-Ledesma ST., Iloilo City Tel. No. 5081045Document24 pages3F Sta. Cruz Bldg. Cor. Fuentes-Ledesma ST., Iloilo City Tel. No. 5081045Chekahay ni 'Cher Ojie ug 'Cher Alven DiazNo ratings yet
- Eye AnatomyDocument21 pagesEye AnatomyHammad afzal100% (2)
- Chapter 7 CELL STRUCTURE AND FUNCTION PDFDocument16 pagesChapter 7 CELL STRUCTURE AND FUNCTION PDFAngelene PelayoNo ratings yet
- Chapter 6 Part 11Document2 pagesChapter 6 Part 11dandymontezNo ratings yet
- Bio Notes 3.3.2 Centriole Flagella, CiliaDocument1 pageBio Notes 3.3.2 Centriole Flagella, CiliaKari Kristine Hoskins BarreraNo ratings yet
- Ciliopatias PDFDocument11 pagesCiliopatias PDFsantiagoluna bNo ratings yet
- Cell Structure and Function: Dr. N. OjehDocument45 pagesCell Structure and Function: Dr. N. OjehZachary HamidNo ratings yet
































































































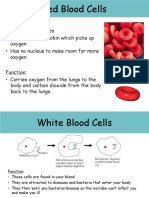







Download as pdf or txt
You might also like
- Cell Modification ModuleDocument30 pagesCell Modification ModuleDuane100% (1)
- USMLE World: Revision QuestionsDocument65 pagesUSMLE World: Revision QuestionsDustin91% (23)
- Phylum Protozoa 02Document25 pagesPhylum Protozoa 02Haris AyubNo ratings yet
- Cilia and FlagellaDocument25 pagesCilia and FlagellaLia Savitri RomdaniNo ratings yet
- Cilia and Flagella: Structure and Function in EukaryotesDocument10 pagesCilia and Flagella: Structure and Function in EukaryotesEr Purushottam PalNo ratings yet
- Science Wps OfficeDocument16 pagesScience Wps OfficeLuke Chase JacintoNo ratings yet
- Locomotion in EuglenaDocument4 pagesLocomotion in Euglenamanojitchatterjee2007No ratings yet
- Locomotary Organelles in ProtozoaDocument25 pagesLocomotary Organelles in Protozoaashley clintonNo ratings yet
- Structure of Cilia and Flagella: - by Neha LoharDocument18 pagesStructure of Cilia and Flagella: - by Neha Loharjoy parimalaNo ratings yet
- Cilia and FlagellaDocument8 pagesCilia and Flagellaamnamalik1304No ratings yet
- Ultrastructure and Development of The Flagellar Apparatus and Flagellar Motion in The Colonial Green Alga AstrephomeneDocument22 pagesUltrastructure and Development of The Flagellar Apparatus and Flagellar Motion in The Colonial Green Alga AstrephomeneARMY SistersNo ratings yet
- Cell Membrane Unit 1-2Document52 pagesCell Membrane Unit 1-2Kemoy FrancisNo ratings yet
- Nihms 367187Document14 pagesNihms 367187Tiego FerreiraNo ratings yet
- Invertebrate Anatomy LengkapDocument49 pagesInvertebrate Anatomy LengkapMayola ArdaNo ratings yet
- Cilia and FlagellaDocument4 pagesCilia and FlagellaAnurag chavan100% (1)
- FlagellumDocument13 pagesFlagellumDaner BaxhijaNo ratings yet
- Structure and FunctionDocument12 pagesStructure and FunctionJustine PamaNo ratings yet
- Semester 1 Locomotion in ProtozoaDocument2 pagesSemester 1 Locomotion in Protozoaashwinirana8No ratings yet
- Microtubules: Organization: Single Microtubules Are Distributed Within The Cytoplasm. Also, MicrotubulesDocument6 pagesMicrotubules: Organization: Single Microtubules Are Distributed Within The Cytoplasm. Also, MicrotubulesArif NabeelNo ratings yet
- Flagellar Restoration in C. ReinhardiDocument8 pagesFlagellar Restoration in C. Reinhardinasek22No ratings yet
- The Cell: Prokaryotic EukaryoticDocument28 pagesThe Cell: Prokaryotic EukaryoticMastersBestHackersNo ratings yet
- The CellDocument24 pagesThe CellCaren AdongoNo ratings yet
- Full Download Zoology 9th Edition Miller Solutions ManualDocument35 pagesFull Download Zoology 9th Edition Miller Solutions Manualcaveneywilliams100% (19)
- 2 PARA 1 - Protozoa - FlagellatesDocument13 pages2 PARA 1 - Protozoa - FlagellatesTricia LlorinNo ratings yet
- Exercise 2: Cells: Examining Cyanobacteria (Blue Green Algae)Document5 pagesExercise 2: Cells: Examining Cyanobacteria (Blue Green Algae)acgonzales28No ratings yet
- Opposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumDocument8 pagesOpposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumFernandoNo ratings yet
- Biol120 Lecture 7Document55 pagesBiol120 Lecture 7mooosemnNo ratings yet
- Lab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeDocument4 pagesLab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeMichael TimsonNo ratings yet
- Morphology and Fine StructureDocument67 pagesMorphology and Fine StructureDiksha PadamNo ratings yet
- The Dynein Family at A GlanceDocument3 pagesThe Dynein Family at A GlanceNovVie VietTha Sccor IINo ratings yet
- EuglenaDocument4 pagesEuglenaBio SciencesNo ratings yet
- Biomed 05Document12 pagesBiomed 05api-3706483No ratings yet
- Module (Cell Nucleus)Document12 pagesModule (Cell Nucleus)Edgie JunelaNo ratings yet
- Parts of A Cell BIOLOGYDocument43 pagesParts of A Cell BIOLOGYSam Ferbyn LomotanNo ratings yet
- Cell and MolecularDocument37 pagesCell and MolecularAbhishek AbhiNo ratings yet
- 1-Ribosomes: Nucleolus Nuclear PoresDocument12 pages1-Ribosomes: Nucleolus Nuclear Poresapi-3706483No ratings yet
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-530699294No ratings yet
- Chapter 2 NeuronDocument49 pagesChapter 2 NeuronAfNo ratings yet
- The Cell: by DR - Hikmat Fatima HashmiDocument53 pagesThe Cell: by DR - Hikmat Fatima Hashmibeverly brittoNo ratings yet
- CellDocument41 pagesCellAditya VermaNo ratings yet
- Cell Unit of Life Question AnswerDocument8 pagesCell Unit of Life Question Answershafiahmad2014.15No ratings yet
- Ultrastructure of Bacteria & EndosporeDocument8 pagesUltrastructure of Bacteria & EndosporeSubrata KunduNo ratings yet
- Kinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardDocument11 pagesKinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardAlessandro BianchessiNo ratings yet
- Elsc Easy Learn Singh Classes: Cell OrganelleDocument13 pagesElsc Easy Learn Singh Classes: Cell OrganelleNandini SinghNo ratings yet
- CH 6 BioDocument5 pagesCH 6 BioedwarddameNo ratings yet
- Cells: H1N3 Publications SDN BHD®Document22 pagesCells: H1N3 Publications SDN BHD®Noor HafizNo ratings yet
- CellsDocument46 pagesCellsErfanShekarrizNo ratings yet
- Organelle: Flagellum DefinitionDocument4 pagesOrganelle: Flagellum DefinitionMadhanNo ratings yet
- Eukaryotic CellsDocument21 pagesEukaryotic Cellsjaysenires13tkdNo ratings yet
- Britannica Print DemoDocument9 pagesBritannica Print DemoduongtuScribdNo ratings yet
- Organelles Working TogetherDocument4 pagesOrganelles Working TogetherVilma Malaya VisarioNo ratings yet
- Assignment II: Macaranas, Mar SDocument7 pagesAssignment II: Macaranas, Mar SMar MacaranasNo ratings yet
- GROUP 5 PhysioLab Activity 3Document7 pagesGROUP 5 PhysioLab Activity 3Deasserei TatelNo ratings yet
- Biochem AssignmentDocument2 pagesBiochem AssignmentAngel GoNo ratings yet
- Lecture 4 Notes 2013Document4 pagesLecture 4 Notes 2013Richard HampsonNo ratings yet
- Cells: A. Ultrastructure & Function of The CellDocument9 pagesCells: A. Ultrastructure & Function of The CellRhea Lyn LamosteNo ratings yet
- Cytoskeleton: Jetro P. MagnayonDocument66 pagesCytoskeleton: Jetro P. MagnayonChristian Santiago100% (1)
- Lab Report 5Document9 pagesLab Report 5jvelezdanielaNo ratings yet
- Cellular LocomotionDocument14 pagesCellular LocomotionEshaNo ratings yet
- Cell Biology Practical 3 PDFDocument11 pagesCell Biology Practical 3 PDFEswaran Singam100% (1)
- Flagella and MotilityDocument17 pagesFlagella and Motilityqasim mughaNo ratings yet
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceFrom EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceNo ratings yet
- Plant Macro-And Micronutrient MineralsDocument5 pagesPlant Macro-And Micronutrient Mineralsmanoj_rkl_07No ratings yet
- Electroporation: Jac A NickoloffDocument3 pagesElectroporation: Jac A Nickoloffmanoj_rkl_07No ratings yet
- DNA Damage: Paul W DoetschDocument7 pagesDNA Damage: Paul W Doetschmanoj_rkl_07No ratings yet
- Phyllosphere PDFDocument8 pagesPhyllosphere PDFmanoj_rkl_07No ratings yet
- Dideoxy Sequencing of DNA PDFDocument16 pagesDideoxy Sequencing of DNA PDFmanoj_rkl_07No ratings yet
- Genetic Code Introduction PDFDocument10 pagesGenetic Code Introduction PDFmanoj_rkl_07No ratings yet
- Closteroviridae: Historical PerspectiveDocument6 pagesClosteroviridae: Historical Perspectivemanoj_rkl_07No ratings yet
- Root Nodules (Rhizobium Legumes) PDFDocument2 pagesRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07No ratings yet
- Terpenoids Lower PDFDocument7 pagesTerpenoids Lower PDFmanoj_rkl_07No ratings yet
- RNA Plant and Animal Virus Replication PDFDocument9 pagesRNA Plant and Animal Virus Replication PDFmanoj_rkl_07No ratings yet
- Plant Tracheary Elements PDFDocument2 pagesPlant Tracheary Elements PDFmanoj_rkl_07No ratings yet
- S Adenosylmethionine PDFDocument7 pagesS Adenosylmethionine PDFmanoj_rkl_07No ratings yet
- Plant Water Relations PDFDocument7 pagesPlant Water Relations PDFmanoj_rkl_07No ratings yet
- Plant Cytoskeleton PDFDocument7 pagesPlant Cytoskeleton PDFmanoj_rkl_07No ratings yet
- Plant Genome Projects PDFDocument4 pagesPlant Genome Projects PDFmanoj_rkl_07100% (1)
- Plant Cells Peroxisomes and Glyoxysomes PDFDocument7 pagesPlant Cells Peroxisomes and Glyoxysomes PDFmanoj_rkl_07No ratings yet
- Cell - The Unit of Life DPP 09 Yakeen NEET 2.0 2025 (LegDocument3 pagesCell - The Unit of Life DPP 09 Yakeen NEET 2.0 2025 (LegBISWARANJAN SAHOONo ratings yet
- Chest Injury and Its TypesDocument14 pagesChest Injury and Its TypesKoochi PoojithaNo ratings yet
- Cell Theory: Biology Scientific Theory CellsDocument5 pagesCell Theory: Biology Scientific Theory CellsNiel S. DefensorNo ratings yet
- Case Studies in Veterinary ImmuneologyDocument209 pagesCase Studies in Veterinary ImmuneologyNathalia gallegoNo ratings yet
- Microbial Morphology and TaxonomyDocument6 pagesMicrobial Morphology and TaxonomyJasmin Pearl AndayaNo ratings yet
- Locomotary Organelles in ProtozoaDocument25 pagesLocomotary Organelles in Protozoaashley clintonNo ratings yet
- Histology and Cell Biology An Introduction To Pathology 5Th Edition Abraham L Kierszenbaum M D PH D Full ChapterDocument67 pagesHistology and Cell Biology An Introduction To Pathology 5Th Edition Abraham L Kierszenbaum M D PH D Full Chapterdennis.roberson462100% (17)
- Cell Organelles - Biology Class 11 - NEETDocument18 pagesCell Organelles - Biology Class 11 - NEETsanjNo ratings yet
- Ws 2 Answer Key Unit 2 22-23Document9 pagesWs 2 Answer Key Unit 2 22-23majdiabd5678No ratings yet
- Test Bank For Color Textbook of Histology 3rd Edition Leslie P GartnerDocument12 pagesTest Bank For Color Textbook of Histology 3rd Edition Leslie P Gartnerbrennaquani2mNo ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- Chapter 6Document13 pagesChapter 6gahaganeNo ratings yet
- Ncert Highlight - Cell - by Seep PahujaDocument18 pagesNcert Highlight - Cell - by Seep PahujaskindustrieshelplineNo ratings yet
- Vol.14 No.3 PDFDocument140 pagesVol.14 No.3 PDFAnn JoseNo ratings yet
- BIOL 102 Lab Manual 2022 - 2023Document184 pagesBIOL 102 Lab Manual 2022 - 2023Saya SinNo ratings yet
- 03 Cell BiologyDocument111 pages03 Cell BiologyPrincess Noreen SavellanoNo ratings yet
- Fragrance Chemistry The Science of The Sense of SmellDocument631 pagesFragrance Chemistry The Science of The Sense of SmellGuedri Ghania0% (1)
- Cell Organelles DefinitionDocument40 pagesCell Organelles Definitionmuhammad aslam0% (1)
- Specialised Cells-& Levels of OrganisationsDocument34 pagesSpecialised Cells-& Levels of OrganisationsEmna MbarekNo ratings yet
- Specialised Cells Information SheetsDocument8 pagesSpecialised Cells Information SheetsVy LyNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- 3F Sta. Cruz Bldg. Cor. Fuentes-Ledesma ST., Iloilo City Tel. No. 5081045Document24 pages3F Sta. Cruz Bldg. Cor. Fuentes-Ledesma ST., Iloilo City Tel. No. 5081045Chekahay ni 'Cher Ojie ug 'Cher Alven DiazNo ratings yet
- Eye AnatomyDocument21 pagesEye AnatomyHammad afzal100% (2)
- Chapter 7 CELL STRUCTURE AND FUNCTION PDFDocument16 pagesChapter 7 CELL STRUCTURE AND FUNCTION PDFAngelene PelayoNo ratings yet
- Chapter 6 Part 11Document2 pagesChapter 6 Part 11dandymontezNo ratings yet
- Bio Notes 3.3.2 Centriole Flagella, CiliaDocument1 pageBio Notes 3.3.2 Centriole Flagella, CiliaKari Kristine Hoskins BarreraNo ratings yet
- Ciliopatias PDFDocument11 pagesCiliopatias PDFsantiagoluna bNo ratings yet
- Cell Structure and Function: Dr. N. OjehDocument45 pagesCell Structure and Function: Dr. N. OjehZachary HamidNo ratings yet