Download as pdf or txt
You might also like
- Project Proposal On Dairy FarmDocument26 pagesProject Proposal On Dairy Farmvector dairy88% (8)
- Broomfield Hospital Site Plan: Café Shops Lifts Information PointsDocument1 pageBroomfield Hospital Site Plan: Café Shops Lifts Information Pointsady trader0% (2)
- ISM CPSM Exam-SpecificationsDocument7 pagesISM CPSM Exam-SpecificationsmsajanjNo ratings yet
- NIH Public Access: Author ManuscriptDocument16 pagesNIH Public Access: Author Manuscript10sgNo ratings yet
- Hal 00893898Document31 pagesHal 00893898Vesee KavezeriNo ratings yet
- Lecture III - Semi-FinalsDocument16 pagesLecture III - Semi-FinalsENGETS IT7No ratings yet
- Struktur Dan Fungsi KromosomDocument32 pagesStruktur Dan Fungsi KromosomAniss MaharaniNo ratings yet
- The Organization of PericentroDocument33 pagesThe Organization of PericentroTunggul AmetungNo ratings yet
- Advance Genetics: THE Complexity of Eukaryotic GenomeDocument9 pagesAdvance Genetics: THE Complexity of Eukaryotic Genomecassidy conchaNo ratings yet
- Yogi - The CellDocument9 pagesYogi - The CellYogi Arta SuarlinNo ratings yet
- In Eukaryotes and Hot PlateDocument7 pagesIn Eukaryotes and Hot PlateNadya Nur TrianaNo ratings yet
- Biologia 1 JoveDocument26 pagesBiologia 1 JoveFernanda NúñezNo ratings yet
- 2 68 1622186738 Ijbrdjun20216Document8 pages2 68 1622186738 Ijbrdjun20216TJPRC PublicationsNo ratings yet
- Structure of Eukaryotic Chromosomes. Karyotype. Methods of Cytogenetic AnalysisDocument4 pagesStructure of Eukaryotic Chromosomes. Karyotype. Methods of Cytogenetic Analysiscarlottabovi28No ratings yet
- Unesco - Eolss Sample Chapters: Evolutionary and Molecular TaxonomyDocument9 pagesUnesco - Eolss Sample Chapters: Evolutionary and Molecular Taxonomycynthiachepkoech343No ratings yet
- Chap. 6 Genes, Genomics, and Chromosomes: TopicsDocument22 pagesChap. 6 Genes, Genomics, and Chromosomes: TopicsSuman AhlyanNo ratings yet
- Chapter 3Document7 pagesChapter 3candra mahardikaNo ratings yet
- Introduction To CytogeneticsDocument40 pagesIntroduction To CytogeneticsJoseph FerreiraNo ratings yet
- Long-Range Organization of Tandem Arrays of A Satellite DNA at The Centromeres of Human ChromosomesDocument5 pagesLong-Range Organization of Tandem Arrays of A Satellite DNA at The Centromeres of Human ChromosomesMariano HorganNo ratings yet
- Nucleosome: Structure and FunctionDocument5 pagesNucleosome: Structure and Functionابراهيم ماهر سلوم حسينNo ratings yet
- Dwi Dutha Arie Sampurna XII A 5 / 08Document18 pagesDwi Dutha Arie Sampurna XII A 5 / 08Dwi Duta AriNo ratings yet
- Eukaryotic, Prokaryotic and Viral Genome OrganisationDocument17 pagesEukaryotic, Prokaryotic and Viral Genome OrganisationAman KhanNo ratings yet
- Lecture 1 - Chromosome & GenomeDocument64 pagesLecture 1 - Chromosome & GenomeFriendlyGoodGirlNo ratings yet
- Poly Chrono Poulos 2014Document12 pagesPoly Chrono Poulos 2014ttz9vvcrrl8gff7006wNo ratings yet
- 1.1 Evolution and Speciation: Chapter OneDocument48 pages1.1 Evolution and Speciation: Chapter OneHassan AL-HillaliNo ratings yet
- GN0303 Notes - Unit1-Part1Document20 pagesGN0303 Notes - Unit1-Part1Abhi KannanNo ratings yet
- Xun Xu The Genomic Sequence Chinese HamsterDocument8 pagesXun Xu The Genomic Sequence Chinese HamsterAbhishek NaikNo ratings yet
- Fransz 2003 ChromR Heterochromatin in Interphase Nuclei of atDocument14 pagesFransz 2003 ChromR Heterochromatin in Interphase Nuclei of atAJMRNo ratings yet
- 12LectF Transcription 2017Document44 pages12LectF Transcription 2017Nofa RanaNo ratings yet
- Genome Organization and Function: A View From Yeast and ArabidopsisDocument13 pagesGenome Organization and Function: A View From Yeast and ArabidopsisJulienne InostrozaNo ratings yet
- Super-Coil Me: Sizing Up Centromeric Nucleosomes: by Emma Hill and Ruth WilliamsDocument4 pagesSuper-Coil Me: Sizing Up Centromeric Nucleosomes: by Emma Hill and Ruth Williams10sgNo ratings yet
- Untitled DocumentDocument21 pagesUntitled Documentakolkarsagar14No ratings yet
- Polytene and Lampbrush ChromosomesDocument3 pagesPolytene and Lampbrush Chromosomesneeru.bhagatNo ratings yet
- Chapter 5 - Basic Concepts in Human Molecular Geneti - 2009 - Molecular PathologDocument19 pagesChapter 5 - Basic Concepts in Human Molecular Geneti - 2009 - Molecular PathologSELNo ratings yet
- Chromosomes WorksheetDocument6 pagesChromosomes WorksheetPaulína PalčinskáNo ratings yet
- Bio101 1Document43 pagesBio101 1usmansulaiman655No ratings yet
- How Genomes EvolveDocument21 pagesHow Genomes EvolveMahmood-S ChoudheryNo ratings yet
- Soma ( Body)Document4 pagesSoma ( Body)Joe AjaNo ratings yet
- Cy To GeneticsDocument5 pagesCy To GeneticsNikoh Anthony EwayanNo ratings yet
- Dna StructureDocument67 pagesDna Structurespitzmark2030No ratings yet
- Proyecto Genoma HumanoDocument50 pagesProyecto Genoma HumanoLila Lubianka Domínguez RamírezNo ratings yet
- Chromosomes: Sardar Hussain Asst - Prof.biotechnology GSC, CtaDocument127 pagesChromosomes: Sardar Hussain Asst - Prof.biotechnology GSC, CtaAshikuzzaman AntorNo ratings yet
- Gene MappingDocument32 pagesGene MappingCLEMENTNo ratings yet
- Human Genome ProjectDocument9 pagesHuman Genome ProjectTanzeela noureenNo ratings yet
- In Brief: Complexity On The NanoscaleDocument1 pageIn Brief: Complexity On The Nanoscaletopgun625No ratings yet
- Unit-I: Characteristics & Structure of GenesDocument64 pagesUnit-I: Characteristics & Structure of GenesMonika JosephNo ratings yet
- A New and Exciting Era of Genomics: No Region Is Beyond ReachDocument2 pagesA New and Exciting Era of Genomics: No Region Is Beyond ReachHaseena SaitNo ratings yet
- C Banding in Mus MusculusDocument10 pagesC Banding in Mus MusculusFlavio DávilaNo ratings yet
- 1 s2.0 S0092867415001324 Main 2Document15 pages1 s2.0 S0092867415001324 Main 2Daff AlfaNo ratings yet
- Plant Centromeric Retrotransposons: A Structural and Cytogenetic PerspectiveDocument16 pagesPlant Centromeric Retrotransposons: A Structural and Cytogenetic PerspectiveFrontiersNo ratings yet
- Plant Genome OrganisationDocument16 pagesPlant Genome OrganisationVerônica D'AddazioNo ratings yet
- Module-1: Genomics and Proteomics NotesDocument8 pagesModule-1: Genomics and Proteomics NotesNishtha KhannaNo ratings yet
- 2011 Koumbaris HumMolGenetDocument12 pages2011 Koumbaris HumMolGenetEduardo PasteneNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument33 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- Content History:: Characteristics of GeneDocument5 pagesContent History:: Characteristics of GeneArchana YadavNo ratings yet
- Structure, Function & Molecular Organization of Eukaryotic ChromosomeDocument18 pagesStructure, Function & Molecular Organization of Eukaryotic Chromosomej_sheriNo ratings yet
- De Novo SATAC Formation 1 Chrom Res 1996Document14 pagesDe Novo SATAC Formation 1 Chrom Res 1996Robert KatonaNo ratings yet
- DNA: The Warehouse/Storage of Genetic Information: by Librada M. RubioDocument16 pagesDNA: The Warehouse/Storage of Genetic Information: by Librada M. RubioAbby SevillaNo ratings yet
- Genes: Centromere Repeats: Hidden Gems of The GenomeDocument22 pagesGenes: Centromere Repeats: Hidden Gems of The Genomebinte farooqNo ratings yet
- Xinwei Shev Et Al - The Structure and Evolution of Centromeric Transition Regions Within The Human GenomeDocument8 pagesXinwei Shev Et Al - The Structure and Evolution of Centromeric Transition Regions Within The Human GenomeYopghm698No ratings yet
- Cell Type AtlasDocument17 pagesCell Type Atlas21lbph02No ratings yet
- Chromosomes: DNA (Deoxyribonucleic Acid) Is The Genetic Material of LivingDocument8 pagesChromosomes: DNA (Deoxyribonucleic Acid) Is The Genetic Material of Livingjoyce sumagaysayNo ratings yet
- 1Document8 pages110sgNo ratings yet
- A Research Agenda For Helminth Diseases of Humans: Modelling For Control and EliminationDocument11 pagesA Research Agenda For Helminth Diseases of Humans: Modelling For Control and Elimination10sgNo ratings yet
- Characterization of Three-Dimensional Spatial Aggregation and Association Patterns of Brown Rot Symptoms Within Intensively Mapped Sour Cherry TreesDocument8 pagesCharacterization of Three-Dimensional Spatial Aggregation and Association Patterns of Brown Rot Symptoms Within Intensively Mapped Sour Cherry Trees10sgNo ratings yet
- Sex-Dependent Novelty Response in Neurexin-1 A Mutant MiceDocument6 pagesSex-Dependent Novelty Response in Neurexin-1 A Mutant Mice10sgNo ratings yet
- Role of Oxidative Stress and The Activity of Ethylene Biosynthetic Enzymes On The Formation of Spongy Tissue in Alphonso' MangoDocument5 pagesRole of Oxidative Stress and The Activity of Ethylene Biosynthetic Enzymes On The Formation of Spongy Tissue in Alphonso' Mango10sgNo ratings yet
- NIH Public Access: Author ManuscriptDocument16 pagesNIH Public Access: Author Manuscript10sgNo ratings yet
- NIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal SyndromeDocument7 pagesNIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal Syndrome10sgNo ratings yet
- Field Attractants For Pachnoda Interrupta Selected by Means of GC-EAD and Single Sensillum ScreeningDocument14 pagesField Attractants For Pachnoda Interrupta Selected by Means of GC-EAD and Single Sensillum Screening10sgNo ratings yet
- Is Axis Position Within Tree Architecture A Determinant of Axis Morphology, Branching, Flowering and Fruiting? An Essay in MangoDocument12 pagesIs Axis Position Within Tree Architecture A Determinant of Axis Morphology, Branching, Flowering and Fruiting? An Essay in Mango10sgNo ratings yet
- Optimization of Serine Protease Purification From Mango (Mangifera Indica Cv. Chokanan) Peel in Polyethylene Glycol/Dextran Aqueous Two Phase SystemDocument14 pagesOptimization of Serine Protease Purification From Mango (Mangifera Indica Cv. Chokanan) Peel in Polyethylene Glycol/Dextran Aqueous Two Phase System10sgNo ratings yet
- Contrasting Diversity Patterns of Crenarchaeal, Bacterial and Fungal Soil Communities in An Alpine LandscapeDocument7 pagesContrasting Diversity Patterns of Crenarchaeal, Bacterial and Fungal Soil Communities in An Alpine Landscape10sgNo ratings yet
- Activation of Ethylene-Responsive P-Hydroxyphenylpyruvate Dioxygenase Leads To Increased Tocopherol Levels During Ripening in MangoDocument11 pagesActivation of Ethylene-Responsive P-Hydroxyphenylpyruvate Dioxygenase Leads To Increased Tocopherol Levels During Ripening in Mango10sgNo ratings yet
- Allergy: Hypersensitivity Manifestations To The Fruit MangoDocument7 pagesAllergy: Hypersensitivity Manifestations To The Fruit Mango10sgNo ratings yet
- Evolution and Organization of A Highly Dynamic, Subtelomeric Helicase Gene Family in The Rice Blast Fungus Magnaporthe GriseaDocument10 pagesEvolution and Organization of A Highly Dynamic, Subtelomeric Helicase Gene Family in The Rice Blast Fungus Magnaporthe Grisea10sgNo ratings yet
- Limits of Nuclear Ribosomal DNA Internal Transcribed Spacer (ITS) Sequences As Species Barcodes For FungiDocument1 pageLimits of Nuclear Ribosomal DNA Internal Transcribed Spacer (ITS) Sequences As Species Barcodes For Fungi10sgNo ratings yet
- The History, Fungal Biodiversity, Conservation, and Future Perspectives For Mycology in EgyptDocument20 pagesThe History, Fungal Biodiversity, Conservation, and Future Perspectives For Mycology in Egypt10sgNo ratings yet
- Marine Drugs: Immense Essence of Excellence: Marine Microbial Bioactive CompoundsDocument29 pagesMarine Drugs: Immense Essence of Excellence: Marine Microbial Bioactive Compounds10sgNo ratings yet
- Soil Eukaryotic Microorganism Succession As Affected by Continuous Cropping of Peanut - Pathogenic and Beneficial Fungi Were SelectedDocument9 pagesSoil Eukaryotic Microorganism Succession As Affected by Continuous Cropping of Peanut - Pathogenic and Beneficial Fungi Were Selected10sgNo ratings yet
- Shifts in Soil Microorganisms in Response To Warming Are Consistent Across A Range of Antarctic EnvironmentsDocument11 pagesShifts in Soil Microorganisms in Response To Warming Are Consistent Across A Range of Antarctic Environments10sgNo ratings yet
- Validation and Application of A PCR Primer Set To Quantify Fungal Communities in The Soil Environment by Real-Time Quantitative PCRDocument13 pagesValidation and Application of A PCR Primer Set To Quantify Fungal Communities in The Soil Environment by Real-Time Quantitative PCR10sgNo ratings yet
- Diazotrophicus On Solid Media To Changes in AtmosphericDocument7 pagesDiazotrophicus On Solid Media To Changes in Atmospheric10sgNo ratings yet
- Relationships of Cereal Crops and Other Grasses: Vol. 95, Pp. 2005-2010, March 1998 Colloquium PaperDocument6 pagesRelationships of Cereal Crops and Other Grasses: Vol. 95, Pp. 2005-2010, March 1998 Colloquium Paper10sgNo ratings yet
- New Strategy For Rapid Diagnosis and Characterization of Fungal Infections: The Example of Corneal ScrapingsDocument10 pagesNew Strategy For Rapid Diagnosis and Characterization of Fungal Infections: The Example of Corneal Scrapings10sgNo ratings yet
- Special Issue Paper: Olorode Afr. J. Trad. CAM (2007) 4 (4) : 532 - 540Document9 pagesSpecial Issue Paper: Olorode Afr. J. Trad. CAM (2007) 4 (4) : 532 - 54010sgNo ratings yet
- Geotechnical Properties of Dublin Boulder ClayDocument18 pagesGeotechnical Properties of Dublin Boulder ClayBLPgalwayNo ratings yet
- 04.02.20 BCTDA MTG Public CommentsDocument85 pages04.02.20 BCTDA MTG Public CommentsDaniel WaltonNo ratings yet
- On Linguistic Politeness Theory: Robin Lakoff's Theory of Politeness, Brown and Levinson's Theory of Politeness, Geoffrey Leech's Theory of PolitenessDocument9 pagesOn Linguistic Politeness Theory: Robin Lakoff's Theory of Politeness, Brown and Levinson's Theory of Politeness, Geoffrey Leech's Theory of PolitenessCentral Asian StudiesNo ratings yet
- Alphard Maintenance PackagesDocument2 pagesAlphard Maintenance PackagesTrader Teknik SqdNo ratings yet
- Mass, Volume, and Density: Goals Shape Formula Symbols Table 2.1Document5 pagesMass, Volume, and Density: Goals Shape Formula Symbols Table 2.1SandyNo ratings yet
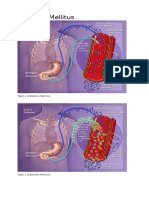
- Acute. KetoacidosisdocxDocument12 pagesAcute. KetoacidosisdocxShara SampangNo ratings yet
- Construct The Relational Model of The Following Scenario. (6 MARKS) System DescriptionDocument2 pagesConstruct The Relational Model of The Following Scenario. (6 MARKS) System DescriptionBushra ShahzadNo ratings yet
- Construction of A 170m Long Cripple Sided Tunnel Using Variable Geometry Hydraulic Formwork in DTL 3, C927Document16 pagesConstruction of A 170m Long Cripple Sided Tunnel Using Variable Geometry Hydraulic Formwork in DTL 3, C927Batu GajahNo ratings yet
- Bluetooth 4.2 SpecificationDocument2,772 pagesBluetooth 4.2 SpecificationScott PooleNo ratings yet
- Um Ugm2006 BinggDocument3 pagesUm Ugm2006 BinggZanaNiswahNabilaNo ratings yet
- Types of Cracks in Concrete BeamsDocument3 pagesTypes of Cracks in Concrete BeamsFatin UmyarahNo ratings yet
- Experiment No. 2: Aim: A) D Flip-Flop: Synchronous VHDL CodeDocument6 pagesExperiment No. 2: Aim: A) D Flip-Flop: Synchronous VHDL CodeRahul MishraNo ratings yet
- Chapter Add On 1Document18 pagesChapter Add On 1Escada BebeyNo ratings yet
- LeetCode 50 Common Interview Questions With Solutions 1654359171Document100 pagesLeetCode 50 Common Interview Questions With Solutions 1654359171Om SharmaNo ratings yet
- Evaluación Manual Del DiafragmaDocument8 pagesEvaluación Manual Del DiafragmaSilvia Victoria SavaroNo ratings yet
- SAP HANA StudioDocument33 pagesSAP HANA StudioPrateekVermaNo ratings yet
- Nami Solar Company Profile Aug 2020 ENGDocument13 pagesNami Solar Company Profile Aug 2020 ENGKien Trung Nguyen100% (1)
- ICC 70 AdministratorsGuide en PDFDocument92 pagesICC 70 AdministratorsGuide en PDFMarie ManuelNo ratings yet
- NLC English 7 Enhancement LP v.1Document85 pagesNLC English 7 Enhancement LP v.1patric maturan100% (1)
- First AId Notes - UpdatedDocument54 pagesFirst AId Notes - UpdatedewawireNo ratings yet
- History of Subcontinent NewspapersDocument1 pageHistory of Subcontinent NewspapersDaniyal Abbas Khan100% (3)
- Perbandingan Jumlah Eritrosit Pada Sampel Darah 3, 2 Dan 1 ML Dengan Antikoagulan K2EdtaDocument6 pagesPerbandingan Jumlah Eritrosit Pada Sampel Darah 3, 2 Dan 1 ML Dengan Antikoagulan K2EdtaNia AzNo ratings yet
- Split Up Syllabus Class Xii Physics 22-23Document1 pageSplit Up Syllabus Class Xii Physics 22-23ROHIT SIHRANo ratings yet
- Abubakar 2014Document13 pagesAbubakar 2014tommy maulanaNo ratings yet
- ThinkPad X1 Carbon 6th Gen Datasheet enDocument3 pagesThinkPad X1 Carbon 6th Gen Datasheet enAnushree SalviNo ratings yet
- Lab Reproductive SDocument8 pagesLab Reproductive SJoyce TorcuatorNo ratings yet
- Thin Film Composite Reverse Osmosis MembranesDocument42 pagesThin Film Composite Reverse Osmosis MembranesGunay Tekkale100% (1)