Download as pdf or txt
You might also like
- Guide To Hazardous Paint Management - ASNZS 4361.2.2017 - Part 2 - Lead Paint in Residential, Public and Commercial BuildingsDocument10 pagesGuide To Hazardous Paint Management - ASNZS 4361.2.2017 - Part 2 - Lead Paint in Residential, Public and Commercial Buildingshitman13630% (1)
- Alex Sander Case StudyDocument12 pagesAlex Sander Case StudyAditya Beri100% (1)
- Critical Review: Structure and Function of VEGF ReceptorsDocument8 pagesCritical Review: Structure and Function of VEGF ReceptorsmonicamoniccNo ratings yet
- Cloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebDocument4 pagesCloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebGJESRNo ratings yet
- Infectious Junin Viruses Differing in Efficient Reverse Genetics Generation ofDocument10 pagesInfectious Junin Viruses Differing in Efficient Reverse Genetics Generation ofLibardo CaraballoNo ratings yet
- Watanabe 2007Document7 pagesWatanabe 2007Abraao Zuza CostaNo ratings yet
- Growth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationDocument28 pagesGrowth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationBf LiawNo ratings yet
- J. Virol.-2003-Sullivan-9733-7Document6 pagesJ. Virol.-2003-Sullivan-9733-7Juan MoralesNo ratings yet
- 2011-Comparative Analysis of Oncogenic Genes RevealedDocument11 pages2011-Comparative Analysis of Oncogenic Genes RevealedASOK KUMARNo ratings yet
- NIH Public Access: Author ManuscriptDocument20 pagesNIH Public Access: Author ManuscriptJason MarshallNo ratings yet
- Research SquareDocument29 pagesResearch SquareMayNeNo ratings yet
- Classification of Infectious Bursal Disease Virus Into GenogroupsDocument10 pagesClassification of Infectious Bursal Disease Virus Into GenogroupsSariSyahruniNo ratings yet
- Inhibition of in Vitro Leukocyte Proliferation byDocument7 pagesInhibition of in Vitro Leukocyte Proliferation byIsrael Espinoza HernándezNo ratings yet
- Auto0711 1335Document13 pagesAuto0711 1335chocosherNo ratings yet
- Neuroscience Research Communications, Vol. 25, NO. 2: (Accepted July 1Document10 pagesNeuroscience Research Communications, Vol. 25, NO. 2: (Accepted July 1Gustav RSRNo ratings yet
- Autophagy Benefits The Replication of Egg Drop Syndrome Virus in Duck Embryo FibroblastsDocument13 pagesAutophagy Benefits The Replication of Egg Drop Syndrome Virus in Duck Embryo FibroblastsmariaNo ratings yet
- Qadeer Et Al 2021Document18 pagesQadeer Et Al 2021Faiz AhmadNo ratings yet
- Journal of Virology - 9557Document6 pagesJournal of Virology - 9557Suryana I. SugandaNo ratings yet
- Expression Profiles of Immune Mediators Infeline Coronavirus-Infected Cells and Clinicalsamples of Feline Coronavirus-Positive CatsDocument13 pagesExpression Profiles of Immune Mediators Infeline Coronavirus-Infected Cells and Clinicalsamples of Feline Coronavirus-Positive CatsRhagaraRhaNo ratings yet
- 2018 Melincovici VegfDocument13 pages2018 Melincovici Vegfdusseldorf27No ratings yet
- Ann Rev of Virology IPNVDocument30 pagesAnn Rev of Virology IPNVRodolfo VelazcoNo ratings yet
- Apoptosis jvb531Document7 pagesApoptosis jvb531Sandra EßbauerNo ratings yet
- European Virology 2000 - Oral AbstractsDocument52 pagesEuropean Virology 2000 - Oral AbstractsElise BalloNo ratings yet
- Jvirol00072 0430Document10 pagesJvirol00072 0430Cipiripi CipiripiNo ratings yet
- Evaluation of A Novel Nested PCR For The RoutineDocument9 pagesEvaluation of A Novel Nested PCR For The RoutineCristina Usuga MonroyNo ratings yet
- tmp88D2 TMPDocument5 pagestmp88D2 TMPFrontiersNo ratings yet
- Morphogenesis of Koi Herpesvirus Observed by Electron MicrosDocument8 pagesMorphogenesis of Koi Herpesvirus Observed by Electron MicrosgiuseppegnrNo ratings yet
- Bahan Paper PatoDocument6 pagesBahan Paper PatoKhairul Ihsan Pengusaha MudaNo ratings yet
- Human HerpesvirusesDocument8 pagesHuman HerpesvirusesevelynseaaNo ratings yet
- Congenital Bleeding DisordersDocument16 pagesCongenital Bleeding Disordersfranciscrick69No ratings yet
- Tagliamonte Et Al., Vaccine 2011Document10 pagesTagliamonte Et Al., Vaccine 2011Luigi BuonaguroNo ratings yet
- Gumboro GenotypeDocument19 pagesGumboro GenotypeSariSyahruniNo ratings yet
- Shekinah1 PDFDocument10 pagesShekinah1 PDFShekinah Andrea DumlaoNo ratings yet
- Complete Genome Sequence of Bovine Viral Diarrhea Virus-1 Strain Egy/Ismailia/2014, Subtype 1bDocument1 pageComplete Genome Sequence of Bovine Viral Diarrhea Virus-1 Strain Egy/Ismailia/2014, Subtype 1bGhada AfifiNo ratings yet
- The ORF8 Protein of SARS-CoV-2 Mediates Immune Evasion Through Potently Downregulating MHC-IDocument41 pagesThe ORF8 Protein of SARS-CoV-2 Mediates Immune Evasion Through Potently Downregulating MHC-IZerohedge71% (7)
- VegfDocument7 pagesVegfBerry BancinNo ratings yet
- Procyanidin Vegf CokDocument12 pagesProcyanidin Vegf CokAmelia SellyNo ratings yet
- Biologicals: SciencedirectDocument9 pagesBiologicals: SciencedirectbrunaNo ratings yet
- Development of Two Competitive Elisas Based On Monoclonal Antibodies For The Serological Detection of Bovine Ephemeral Fever VirusDocument16 pagesDevelopment of Two Competitive Elisas Based On Monoclonal Antibodies For The Serological Detection of Bovine Ephemeral Fever VirusJovanaNo ratings yet
- Hemorrhagic and Clotting Abnormalities in Infectious Bursal Disease in Specific-Pathogen-Free ChicksDocument8 pagesHemorrhagic and Clotting Abnormalities in Infectious Bursal Disease in Specific-Pathogen-Free ChicksHikmat UllahNo ratings yet
- 08 Coronavirus Felino 2012Document8 pages08 Coronavirus Felino 2012fitoNo ratings yet
- Retrovirus Producer Cells Encoding Antisense Vegf Prolong Survival of Rats With Intracranial GS9L GliomasDocument14 pagesRetrovirus Producer Cells Encoding Antisense Vegf Prolong Survival of Rats With Intracranial GS9L GliomasDong DongNo ratings yet
- Dengue Fever Disease ProcessDocument12 pagesDengue Fever Disease ProcessJake PaduaNo ratings yet
- Vaccines 09 00343Document26 pagesVaccines 09 00343scolopaxNo ratings yet
- 2020 Distinct Infection Process of SARS CoV 2 in Human Bronchial Epithelial Cell LinesDocument9 pages2020 Distinct Infection Process of SARS CoV 2 in Human Bronchial Epithelial Cell LinesMartin JYNo ratings yet
- Molecular Characterization of The Isolated Strains of Bovine Viral Diarrhea VirusDocument7 pagesMolecular Characterization of The Isolated Strains of Bovine Viral Diarrhea VirusGhada AfifiNo ratings yet
- Veterinary Pathology Online: Equine Viral ArteritisDocument11 pagesVeterinary Pathology Online: Equine Viral ArteritismeawchiNo ratings yet
- 蜱唾液肽HIDfsin2通过增强p38信号通路在体外促进蜱传病毒SFTSV复制Document12 pages蜱唾液肽HIDfsin2通过增强p38信号通路在体外促进蜱传病毒SFTSV复制HAOXIANG ZHANGNo ratings yet
- Review 2015 EBV Glycoproteins - Where Are We NowDocument13 pagesReview 2015 EBV Glycoproteins - Where Are We Nowkha diNo ratings yet
- Insect Cell CultureDocument6 pagesInsect Cell CultureJakobsen2012No ratings yet
- Classical Swine Fever-An Updated ReviewDocument25 pagesClassical Swine Fever-An Updated ReviewNatalia PatiñoNo ratings yet
- Classical Swine Fever-An Updated ReviewDocument25 pagesClassical Swine Fever-An Updated ReviewNatalia PatiñoNo ratings yet
- AG110 Is A Novel, Highly Selective Inhibitor The Imidazopyrrolopyridine AnalogueDocument9 pagesAG110 Is A Novel, Highly Selective Inhibitor The Imidazopyrrolopyridine Analoguemariale00veNo ratings yet
- Signatures in Sars-Cov-2 Spike Protein Conferring Escape To Neutralizing AntibodiesDocument25 pagesSignatures in Sars-Cov-2 Spike Protein Conferring Escape To Neutralizing AntibodiesrezqiNo ratings yet
- EGFR Transactivation Contributes To Neuroinflammation in Streptococcus Suis MeningitisDocument14 pagesEGFR Transactivation Contributes To Neuroinflammation in Streptococcus Suis MeningitisBlink SNo ratings yet
- ZD6474 Inhibits Vascular Endothelial Growth Factor Signaling, Angiogenesis, and Tumor Growth Following Oral AdministrationDocument11 pagesZD6474 Inhibits Vascular Endothelial Growth Factor Signaling, Angiogenesis, and Tumor Growth Following Oral AdministrationGregory PeckNo ratings yet
- Newcastle Disease Virus Detection From Chicken Organ Samples Using Reverse Transcriptase Polymerase Chain ReactionDocument14 pagesNewcastle Disease Virus Detection From Chicken Organ Samples Using Reverse Transcriptase Polymerase Chain ReactionKasmira miraNo ratings yet
- Reviewer DairyDocument43 pagesReviewer DairyLandcel Clarence ArcedoNo ratings yet
- Whole Genome Sequencing of Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV) From Field Clinical Samples Improves The Genomic Surveillance of The VirusDocument36 pagesWhole Genome Sequencing of Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV) From Field Clinical Samples Improves The Genomic Surveillance of The VirusJephte LaputNo ratings yet
- Ebola Virus-Like Particles Protect From Lethal Ebola Virus InfectionDocument6 pagesEbola Virus-Like Particles Protect From Lethal Ebola Virus InfectionZeuqzav RzNo ratings yet
- Recombinant DNA TechnologyDocument66 pagesRecombinant DNA TechnologySrishti SikariaNo ratings yet
- 2000-Merwin & BrundigeDocument8 pages2000-Merwin & BrundigeMestesanu ElisabetaNo ratings yet
- Serologic AND Experimental Investigations OF Contagious Ecthyma in AlaskaDocument5 pagesSerologic AND Experimental Investigations OF Contagious Ecthyma in AlaskaMestesanu ElisabetaNo ratings yet
- Poxviridae: In: A Concise Review of Veterinary Virology, Carter G.R., Wise D.J. and Flores E.F. (Eds.) - A3410.1005Document10 pagesPoxviridae: In: A Concise Review of Veterinary Virology, Carter G.R., Wise D.J. and Flores E.F. (Eds.) - A3410.1005Mestesanu ElisabetaNo ratings yet
- Free Pattern DLDocument2 pagesFree Pattern DLMestesanu ElisabetaNo ratings yet
- Biology Shapta Assessment 1 2023Document8 pagesBiology Shapta Assessment 1 2023Nsaiga RonaldNo ratings yet
- International Journal of Disaster Risk Reduction: Lyca G. Ipong, Elvira E. Ongy, Milagros C. BalesDocument9 pagesInternational Journal of Disaster Risk Reduction: Lyca G. Ipong, Elvira E. Ongy, Milagros C. BalesChristian Jeremy ManaloNo ratings yet
- Four Handed DentistryDocument114 pagesFour Handed DentistryLie FelixNo ratings yet
- Dalair LimitedHealthSafetyManualDocument25 pagesDalair LimitedHealthSafetyManualRidarto LubisNo ratings yet
- Aab e Rehmat New Plant Installation Proposal - FY2021-22Document9 pagesAab e Rehmat New Plant Installation Proposal - FY2021-22Amna W. KhalidNo ratings yet
- An Orgasm Is Who Defines What An Orgasm Is?: Sexual and Relationship TherapyDocument8 pagesAn Orgasm Is Who Defines What An Orgasm Is?: Sexual and Relationship Therapytharuka1000No ratings yet
- CP05-03-MMC-MST-ELE-0027 REV.00 Testing & Commissioning For Emergency Lighting SystemDocument35 pagesCP05-03-MMC-MST-ELE-0027 REV.00 Testing & Commissioning For Emergency Lighting SystemMohamed AshrafNo ratings yet
- Metro Manila: Las Piñas City of Manila Parañaque PaterosDocument19 pagesMetro Manila: Las Piñas City of Manila Parañaque PaterosMEDIPAD SMKCI-CABAGANNo ratings yet
- Material Safety Data Sheet: Section 1: Product and Company IdentificationDocument7 pagesMaterial Safety Data Sheet: Section 1: Product and Company IdentificationJUAN GUTIERREZNo ratings yet
- THERAPEUTIC Nurse Patient RELATIONSHIPDocument68 pagesTHERAPEUTIC Nurse Patient RELATIONSHIPDhAiRyA ArOrA91% (11)
- QuestionnaireDocument4 pagesQuestionnaireIan AtienzaNo ratings yet
- EVS PPT Air PollutionDocument27 pagesEVS PPT Air PollutionKajal SainiNo ratings yet
- q2 Health - 8 M 4 Revised PDFDocument24 pagesq2 Health - 8 M 4 Revised PDFGemarie CallosaNo ratings yet
- Heart Attack Detection and AlerDocument3 pagesHeart Attack Detection and Alerprabhujaya97893No ratings yet
- Chapter 2Document2 pagesChapter 2Reuel Gian SadangNo ratings yet
- SDS Canada EU English - Uvex Clear Solution - Honeywell4 - S471 S482 S483 S484Document7 pagesSDS Canada EU English - Uvex Clear Solution - Honeywell4 - S471 S482 S483 S484Henry1249No ratings yet
- Viton GFDocument36 pagesViton GFAlejandro ZagalNo ratings yet
- Normal Value For Urine TestDocument2 pagesNormal Value For Urine TestMarcellino NernereNo ratings yet
- Kav It HaDocument158 pagesKav It HaRuthraaadi RameshkumarNo ratings yet
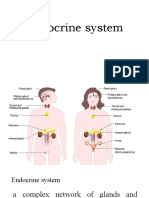
- Endocrine SystemDocument12 pagesEndocrine SystemGiselle AlindoganNo ratings yet
- Stoller Alpha-1AntitrypsinDeficiency 2017Document8 pagesStoller Alpha-1AntitrypsinDeficiency 2017FrancoNo ratings yet
- Pro6 Infotext: Hotline Numbers: Smart 0949 883 3741 0921 939 3004 Globe 0917 594 3753Document2 pagesPro6 Infotext: Hotline Numbers: Smart 0949 883 3741 0921 939 3004 Globe 0917 594 3753Deo Quimpo QuinicioNo ratings yet
- Technical Committee On ExplosivesDocument34 pagesTechnical Committee On Explosivesrusli bahtiarNo ratings yet
- Airway DevicesDocument58 pagesAirway DevicesArmaanjeet SinghNo ratings yet
- ENGLISH 9th GradeDocument17 pagesENGLISH 9th GradeMario SuárezNo ratings yet
- Digital Detox - WorksheetDocument12 pagesDigital Detox - WorksheetFabiana Alejandra RuizNo ratings yet
- Communication Skills For Children With Severe Learning DifficultiesDocument10 pagesCommunication Skills For Children With Severe Learning DifficultiesMay Grace E. BalbinNo ratings yet
- Menstural Taboos in NepalDocument14 pagesMenstural Taboos in Nepalbijaya08No ratings yet