Download as pdf or txt
You might also like
- Mapeh 10 - Third-Quarter-ExamDocument4 pagesMapeh 10 - Third-Quarter-ExamGerome Zamora77% (22)
- Structured Clinical Interview For DSM-IV Axis I DisordersDocument223 pagesStructured Clinical Interview For DSM-IV Axis I DisordersΕιρηνη Κουτσοδοντη100% (2)
- Samer S. Hoz (Auth.) - Vascular Neurosurgery - in Multiple-Choice Questions (2017, Springer International Publishing) PDFDocument223 pagesSamer S. Hoz (Auth.) - Vascular Neurosurgery - in Multiple-Choice Questions (2017, Springer International Publishing) PDFdedi100% (1)
- Culture-Dependent and Culture-IndependenDocument7 pagesCulture-Dependent and Culture-Independenじょしら フィアンナNo ratings yet
- Infection With Koi Herpesvirus: 1. ScopeDocument18 pagesInfection With Koi Herpesvirus: 1. ScopeVira Rizqi RahmawatiNo ratings yet
- Research Article: Role of Flies As Vectors of Foodborne Pathogens in Rural AreasDocument8 pagesResearch Article: Role of Flies As Vectors of Foodborne Pathogens in Rural AreasTércio MeloNo ratings yet
- Experimental ParasitologyDocument4 pagesExperimental ParasitologyShellGarciaNo ratings yet
- Virus ResearchDocument6 pagesVirus ResearchDiego TulcanNo ratings yet
- Varroa Mites and Honey Bee Health Can Varroa Explain PartDocument11 pagesVarroa Mites and Honey Bee Health Can Varroa Explain PartMelivea Paez HerediaNo ratings yet
- Susceptibility of Nile Tilapia (Oreochromis Niloticus) To Vibriosis Due To Vibrio Vulnificus Biotype 2 (Serovar E)Document10 pagesSusceptibility of Nile Tilapia (Oreochromis Niloticus) To Vibriosis Due To Vibrio Vulnificus Biotype 2 (Serovar E)Gianela Judith Vega PinedoNo ratings yet
- Comparison of Mortality and Viral Load in Rainbow Trout G1 y G5Document8 pagesComparison of Mortality and Viral Load in Rainbow Trout G1 y G5Rodolfo VelazcoNo ratings yet
- Arboviruses: Robert E. Shope and James M. MeeganDocument2 pagesArboviruses: Robert E. Shope and James M. Meeganfelipe andreNo ratings yet
- Cẩm Nang Nuôi Dạy ChóDocument4 pagesCẩm Nang Nuôi Dạy ChóĐịnh PhanNo ratings yet
- 2011 PerinottoDocument4 pages2011 PerinottoYenia Micol MatwiejukNo ratings yet
- Ecological Divergence of Two Sympatric LDocument12 pagesEcological Divergence of Two Sympatric LHussein KHNo ratings yet
- Fontana 2020Document9 pagesFontana 2020JUDITH ENIT ROLDAN RODRIGUEZNo ratings yet
- The Problem and Its ScopeDocument4 pagesThe Problem and Its ScopeCJ ManlangitNo ratings yet
- 436 2010 Article 2064Document6 pages436 2010 Article 2064Arif SetiawanNo ratings yet
- Warmer Temperatures Reduce The Vectorial Capacity of Malaria MosquitoesDocument4 pagesWarmer Temperatures Reduce The Vectorial Capacity of Malaria Mosquitoesibrahima1968No ratings yet
- Shiva Chandra 2011Document16 pagesShiva Chandra 2011Spica RegulusNo ratings yet
- White Nose SyndromeDocument25 pagesWhite Nose SyndromeSaifuddin HaswareNo ratings yet
- Microbial Infection Affects Egg Viability and Incubation Behavior in A Tropical PasserineDocument7 pagesMicrobial Infection Affects Egg Viability and Incubation Behavior in A Tropical PasserineMohammed KouidriNo ratings yet
- Ogdenetal JME2004Document13 pagesOgdenetal JME2004Irina AtudoreiNo ratings yet
- CatfluDocument13 pagesCatflumvdaniela.santanaNo ratings yet
- CH#04Document12 pagesCH#04Emran khanNo ratings yet
- Cronobacter (Enterobacter Sakazakii)Document12 pagesCronobacter (Enterobacter Sakazakii)12CH1-13 ĐỖ HOÀNG PHÚCNo ratings yet
- 2010 Healy Et Al. - Review, Cronobacter (Enterobacter Sakazakii)Document15 pages2010 Healy Et Al. - Review, Cronobacter (Enterobacter Sakazakii)Yesica Marcelina SinagaNo ratings yet
- TJV 014Document7 pagesTJV 014Nhe FirmansyahNo ratings yet
- Souza-Neto Powell Bonizzoni. 2019. Aedes Aegypti Vector Competence Studies. A Review.Document19 pagesSouza-Neto Powell Bonizzoni. 2019. Aedes Aegypti Vector Competence Studies. A Review.Laura FloresNo ratings yet
- DrosDocument15 pagesDrosAna María OchoaNo ratings yet
- Egyptian Journal of Aquatic Research: Eman El-Wazzan, Doaa A. Ghareeb, Bahaa AbdellaDocument6 pagesEgyptian Journal of Aquatic Research: Eman El-Wazzan, Doaa A. Ghareeb, Bahaa Abdellazaenal abidinNo ratings yet
- 34 PDFDocument9 pages34 PDFYovan BlancoNo ratings yet
- Effects of Different PH Levels On The Viability, Metamorphosis Rate and Morphology of Aedes MosquitoesDocument28 pagesEffects of Different PH Levels On The Viability, Metamorphosis Rate and Morphology of Aedes Mosquitoesamagno7891No ratings yet
- Article 1 SpirocercaDocument33 pagesArticle 1 SpirocercaJuan Vera SanchezNo ratings yet
- J TVJL 2007 11 012Document14 pagesJ TVJL 2007 11 012ewNo ratings yet
- International Journal For Parasitology: Katja Fischer, Shelley WaltonDocument13 pagesInternational Journal For Parasitology: Katja Fischer, Shelley Waltonduverney.gaviriaNo ratings yet
- Environmental Persistence of Amphibian and Reptilian RanavirusesDocument8 pagesEnvironmental Persistence of Amphibian and Reptilian RanavirusesJamesComeyJustaBitchNo ratings yet
- Chapitre Abalone HerpesvirusDocument12 pagesChapitre Abalone HerpesvirusmiftahuddinmadjidNo ratings yet
- 16 Peralta-Sanchez 2012 IBISDocument13 pages16 Peralta-Sanchez 2012 IBISalcedo76No ratings yet
- A Research For An Eco Friendly Mosquito Control by Using A New Mosquito Trap Mos Hole Trap in A Cowshed in Yeoju of KoreaDocument10 pagesA Research For An Eco Friendly Mosquito Control by Using A New Mosquito Trap Mos Hole Trap in A Cowshed in Yeoju of KoreaJasmin Avila VargasNo ratings yet
- Diptera e ContaminaçãoDocument24 pagesDiptera e Contaminaçãogustavo paimNo ratings yet
- Host Specificity, Pathogen Exposure, and Superinfections Impact TheDocument14 pagesHost Specificity, Pathogen Exposure, and Superinfections Impact TheSarai MontielNo ratings yet
- Solignat Et Al. - 2009 - Replication Cycle of Chikungunya A Re-Emerging ArbovirusDocument15 pagesSolignat Et Al. - 2009 - Replication Cycle of Chikungunya A Re-Emerging ArbovirusLaise Eduarda Paixão de MoraesNo ratings yet
- Phage Therapy: The Escherichia Coli Experience: ReviewDocument8 pagesPhage Therapy: The Escherichia Coli Experience: ReviewAbhay Kumar MohiniNo ratings yet
- Bài 17 - Prevalence of Histopathological Intestinal Lesions and Enteric Pathogens in Dutch Commercial Broilers With TimeDocument11 pagesBài 17 - Prevalence of Histopathological Intestinal Lesions and Enteric Pathogens in Dutch Commercial Broilers With Time21112309No ratings yet
- Porcine Respiratory Coronavirus: Molecular Features and Virus-Host InteractionsDocument27 pagesPorcine Respiratory Coronavirus: Molecular Features and Virus-Host InteractionsTratorbras ZanattaNo ratings yet
- 9e58 PDFDocument10 pages9e58 PDFcarlos javierNo ratings yet
- Journal of Virology-2010-Graham-3134.fullDocument13 pagesJournal of Virology-2010-Graham-3134.fullJohn WilliamsonNo ratings yet
- Micro IngDocument7 pagesMicro IngValentina EspinosaNo ratings yet
- Nature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetenceDocument30 pagesNature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetencedianamfgNo ratings yet
- 1 s2.0 S1567134821004937 MainDocument9 pages1 s2.0 S1567134821004937 Mainchristina kusuma reniNo ratings yet
- Aphid Transmission of The Ontario Isolate of PlumDocument7 pagesAphid Transmission of The Ontario Isolate of PlumDamian PiglionicoNo ratings yet
- Lezama-Gutiérrez Et Al. - 2005 - Susceptibility of Biological Stages of The Horn FLDocument8 pagesLezama-Gutiérrez Et Al. - 2005 - Susceptibility of Biological Stages of The Horn FLJuan UrrutiaNo ratings yet
- Toxoplasma Gondii Infection in Philippines Rattus Spp. Confi RmedDocument12 pagesToxoplasma Gondii Infection in Philippines Rattus Spp. Confi RmedHassanNo ratings yet
- Comparative Studies of Nairobi-Sheep-Disease Virus Strains Infectivity, Immunogenicty and Cross- Protectivity in BALB/C Mice ModelDocument8 pagesComparative Studies of Nairobi-Sheep-Disease Virus Strains Infectivity, Immunogenicty and Cross- Protectivity in BALB/C Mice ModelInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- HFPCH01Document22 pagesHFPCH01collegeassignmenthelp813No ratings yet
- Small Mosquitoes, Large Implications: Crowding and Starvation Affects Gene Expression and Nutrient Accumulation in Aedes AegyptiDocument14 pagesSmall Mosquitoes, Large Implications: Crowding and Starvation Affects Gene Expression and Nutrient Accumulation in Aedes AegyptiMahmoud el gendyNo ratings yet
- Relation to Enterocins and Herbal Extracts of Fecal Hemolytic Es 2019 PoultrDocument7 pagesRelation to Enterocins and Herbal Extracts of Fecal Hemolytic Es 2019 PoultrAleneNo ratings yet
- Understanding The Biological and Molecular Dynamics of Emarging PandemicDocument20 pagesUnderstanding The Biological and Molecular Dynamics of Emarging PandemicFavour OgbonnaNo ratings yet
- Prevalence of NewcastleDocument8 pagesPrevalence of NewcastleDanladi Idris HassanNo ratings yet
- E Coli fnx050Document11 pagesE Coli fnx050Angie LeeNo ratings yet
- Alphavirus Identification in Neotropical BatsDocument11 pagesAlphavirus Identification in Neotropical BatsItrux jonathan cisnerosNo ratings yet
- Bioelectricity: Section 1 - Make PlansDocument71 pagesBioelectricity: Section 1 - Make PlansAnirudh AcharyaNo ratings yet
- Pouch and Will Develop Into The Anterior Pituitary orDocument130 pagesPouch and Will Develop Into The Anterior Pituitary orAnirudh AcharyaNo ratings yet
- Population and Parity Levels of Aedes Aegypti Collected in TucsonDocument9 pagesPopulation and Parity Levels of Aedes Aegypti Collected in TucsonAnirudh AcharyaNo ratings yet
- Guidelines: Food and Agriculture Organization of The United NationsDocument16 pagesGuidelines: Food and Agriculture Organization of The United NationsAnirudh AcharyaNo ratings yet
- Trend of Malaria in KarnatakaDocument8 pagesTrend of Malaria in KarnatakaAnirudh AcharyaNo ratings yet
- Hypothalamus and Its Hormones, Hormones of The Pituitary GlandDocument28 pagesHypothalamus and Its Hormones, Hormones of The Pituitary GlandAnirudh AcharyaNo ratings yet
- Chapter 3 Recombinant DNA TechnologyDocument39 pagesChapter 3 Recombinant DNA TechnologyAnirudh Acharya50% (2)
- Western TragopanDocument5 pagesWestern TragopanAnirudh AcharyaNo ratings yet
- CephalopodDocument14 pagesCephalopodAnirudh AcharyaNo ratings yet
- Neuroendocrine of CrustaceaDocument29 pagesNeuroendocrine of CrustaceaAnirudh Acharya100% (1)
- PomcDocument8 pagesPomcAnirudh AcharyaNo ratings yet
- Ames TestDocument16 pagesAmes TestAnirudh AcharyaNo ratings yet
- SI No. Date Experiments Page NoDocument1 pageSI No. Date Experiments Page NoAnirudh AcharyaNo ratings yet
- Pests of CerealsDocument22 pagesPests of CerealsAnirudh AcharyaNo ratings yet
- Presented By:: Anirudh R. Acharya, M.SC - II Semester, Department of Zoology, University of MysoreDocument38 pagesPresented By:: Anirudh R. Acharya, M.SC - II Semester, Department of Zoology, University of MysoreAnirudh AcharyaNo ratings yet
- 402 Exam Notes 402 Exam NotesDocument13 pages402 Exam Notes 402 Exam NotesMSNo ratings yet
- 7 GEN ED Upon Seeing Water Flowing and FloodingDocument12 pages7 GEN ED Upon Seeing Water Flowing and FloodingLeih Anne Quezon AnonuevoNo ratings yet
- Program Faculty - Sports MedicineDocument17 pagesProgram Faculty - Sports MedicineKavirNivNo ratings yet
- Times Leader 01-14-2012Document46 pagesTimes Leader 01-14-2012The Times LeaderNo ratings yet
- Republic Act No. 10707 Probation Law As Amended PD 968Document20 pagesRepublic Act No. 10707 Probation Law As Amended PD 968Leopardas Lassang CarlosNo ratings yet
- PsychotherapyDocument4 pagesPsychotherapyFaustrock Dionyso MingonkNo ratings yet
- Branched Neurology ExamDocument5 pagesBranched Neurology ExamJacqueline LongNo ratings yet
- Ostomy Care in Home Health1Document39 pagesOstomy Care in Home Health1api-315120220100% (1)
- Hook Surgery Practice Booklet PDFDocument4 pagesHook Surgery Practice Booklet PDFAnonymous b9RIEMekNo ratings yet
- Basic First AidDocument81 pagesBasic First AidDrVarun Menon100% (1)
- Data DictionaryDocument5 pagesData DictionaryDavid JosephNo ratings yet
- Wound DressingDocument18 pagesWound DressingBabita DhruwNo ratings yet
- Hubungan Pelaksanaan Screening Test Menelan Dengan Kejadian Disfagia Pada Pasien Baru Yang Menderita Stroke AkutDocument7 pagesHubungan Pelaksanaan Screening Test Menelan Dengan Kejadian Disfagia Pada Pasien Baru Yang Menderita Stroke AkutmaydamayNo ratings yet
- Control of The Vertical DimensionDocument10 pagesControl of The Vertical DimensionThang Nguyen TienNo ratings yet
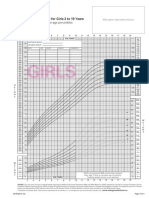
- Girls: WHO Set 2 Growth Chart For Girls 2 To 19 YearsDocument2 pagesGirls: WHO Set 2 Growth Chart For Girls 2 To 19 YearseuiskurNo ratings yet
- Nurs 605 Week 5 PNSG TableDocument16 pagesNurs 605 Week 5 PNSG Tableapi-429205355No ratings yet
- Bronchopneumonia PP 2003Document31 pagesBronchopneumonia PP 2003Rizky FadhilaNo ratings yet
- Fertility InnDocument15 pagesFertility Innkunnamon1234100% (2)
- Acid FAst BacilliDocument6 pagesAcid FAst BacilliGd ManikNo ratings yet
- Rules For Scoring Respiratory Events in Sleep: Update of The 2007 AASM Manual For The Scoring of Sleep and Associated EventsDocument23 pagesRules For Scoring Respiratory Events in Sleep: Update of The 2007 AASM Manual For The Scoring of Sleep and Associated Eventsgiulio pinnolaNo ratings yet
- Yoga Poses and What They AchieveDocument1 pageYoga Poses and What They AchieveVenketeshwar SwamiNo ratings yet
- Anat100 Tutorial 1Document2 pagesAnat100 Tutorial 1Teni LawalNo ratings yet
- R-Peak Detection Algorithm For Ecg Using Double Difference and RR Interval ProcessingDocument5 pagesR-Peak Detection Algorithm For Ecg Using Double Difference and RR Interval ProcessingArash MinNo ratings yet
- Dravya Guna Shastra - The Knowledge of The Qualities of SubstancesDocument12 pagesDravya Guna Shastra - The Knowledge of The Qualities of SubstancesSN WijesinheNo ratings yet
- 2018 Postgraduate 3110Document32 pages2018 Postgraduate 3110Mi XieNo ratings yet
- How To Recoil A Vape CoilDocument11 pagesHow To Recoil A Vape CoilAlldelectro musicNo ratings yet
- Hilot: (Wellness Massage) NC IiDocument44 pagesHilot: (Wellness Massage) NC IiMechel PurcaNo ratings yet