Download as pdf or txt
You might also like
- Hitech Horticulture PDFDocument7 pagesHitech Horticulture PDFSofi MehrajNo ratings yet
- PHB Production in Azomonas, Acinteobacter and Bacillus Species: Isolation, Screening and IdentificationDocument8 pagesPHB Production in Azomonas, Acinteobacter and Bacillus Species: Isolation, Screening and IdentificationInternational Medical PublisherNo ratings yet
- Contrera Et Al 2013 NMRDocument10 pagesContrera Et Al 2013 NMRSurajit BhattacharjeeNo ratings yet
- Biotech Roject 1Document30 pagesBiotech Roject 1bhaskar VermaNo ratings yet
- Biodegradable Plastic A Potential Substi PDFDocument8 pagesBiodegradable Plastic A Potential Substi PDFBrynt Yuan Lord PlacidesNo ratings yet
- Brazilian Journal of MicrobiologyDocument7 pagesBrazilian Journal of MicrobiologyHimaniNo ratings yet
- Optimization of Cellulase Enzyme From Vegetable Waste by Using Trichoderma Atroviride in Solid State FermentationDocument6 pagesOptimization of Cellulase Enzyme From Vegetable Waste by Using Trichoderma Atroviride in Solid State FermentationIOSRjournalNo ratings yet
- Production and Optimization of Bioplastic From Bacillus Isolated From Environmental SampleDocument13 pagesProduction and Optimization of Bioplastic From Bacillus Isolated From Environmental SampleIJAR JOURNALNo ratings yet
- Bioconversion of Cassava Starch By-Product Into Bacillus and Related Bacteria PolyhydroxyalkanoatesDocument13 pagesBioconversion of Cassava Starch By-Product Into Bacillus and Related Bacteria PolyhydroxyalkanoatesMarelin Pereda SifuentesNo ratings yet
- Valorization of Ibombax CeibDocument23 pagesValorization of Ibombax CeibAbdullah Nur HakamiNo ratings yet
- Optimization of Bio-Mosquitocidal Production Using Local Bacterial Strains of Bacillus SphaericusDocument11 pagesOptimization of Bio-Mosquitocidal Production Using Local Bacterial Strains of Bacillus SphaericusTejas DoshiNo ratings yet
- Accepted Manuscript: Bioresource TechnologyDocument18 pagesAccepted Manuscript: Bioresource TechnologyRuxandra BadiuNo ratings yet
- Rodrguez Contreras 2013Document10 pagesRodrguez Contreras 2013fernandoNo ratings yet
- Freitas2018 PDFDocument11 pagesFreitas2018 PDFVanesa CaroNo ratings yet
- The Study of Shelf Life For Liquid Biofertilizer From Vegetable WasteDocument6 pagesThe Study of Shelf Life For Liquid Biofertilizer From Vegetable WasteFadhilah SurotoNo ratings yet
- Welcome To International Journal of Engineering Research and Development (IJERD)Document6 pagesWelcome To International Journal of Engineering Research and Development (IJERD)IJERDNo ratings yet
- Dubrovskis 2007 Biogas ZapalloDocument10 pagesDubrovskis 2007 Biogas ZapalloJulio ReyesNo ratings yet
- Bioplastics RRLDocument16 pagesBioplastics RRLAbegail Olivar100% (1)
- Bacterias - B.megateriumDocument7 pagesBacterias - B.megateriumCatalinaManjarresNo ratings yet
- FPD679.pdf Version 1Document6 pagesFPD679.pdf Version 1Ali IrtazaNo ratings yet
- 5580976Document7 pages5580976Sura C. JirNo ratings yet
- Ashah 2020 IOP Conf. Ser. Earth Environ. Sci. 476 012105Document9 pagesAshah 2020 IOP Conf. Ser. Earth Environ. Sci. 476 012105HarshNo ratings yet
- Bioresource Technology: Ho Myeong Kim, Chi Hoon Oh, Hyeun-Jong BaeDocument7 pagesBioresource Technology: Ho Myeong Kim, Chi Hoon Oh, Hyeun-Jong BaeLaura Ximena Vargas GutiérrezNo ratings yet
- Appl. Environ. Microbiol. 2009 Ibrahim 6222 31Document10 pagesAppl. Environ. Microbiol. 2009 Ibrahim 6222 31ashuna249396No ratings yet
- PHB Paper IJMRT March 2015 Copy1Document11 pagesPHB Paper IJMRT March 2015 Copy1Arjun JayakumarNo ratings yet
- 1 PBDocument6 pages1 PBjaka djNo ratings yet
- Aljuboori Et Al.. 2014. Production of A Bioflocculant From Aspergillus Niger Using Palm Oil Mill Effluent As Carbon SourceDocument5 pagesAljuboori Et Al.. 2014. Production of A Bioflocculant From Aspergillus Niger Using Palm Oil Mill Effluent As Carbon SourceAzb 711No ratings yet
- 1 s2.0 S2214785316303583 MainDocument6 pages1 s2.0 S2214785316303583 MainMaría Camila GonzalezNo ratings yet
- Bajaj, Singhal. 2011. Flocculation Properties of Poly (γ-Glutamic Acid) Produced from Bacillus subtilis IsolateDocument8 pagesBajaj, Singhal. 2011. Flocculation Properties of Poly (γ-Glutamic Acid) Produced from Bacillus subtilis IsolateAzb 711No ratings yet
- Microbiological and Physico-Chemical Dynamics During The Fermentation of The Millet-Based and The Sorghum-Based in The Republic of BeninDocument9 pagesMicrobiological and Physico-Chemical Dynamics During The Fermentation of The Millet-Based and The Sorghum-Based in The Republic of BeninOpenaccess Research paperNo ratings yet
- 1 s2.0 S0956053X18305063 MainDocument7 pages1 s2.0 S0956053X18305063 MainRick Anthony Rosal JaramilloNo ratings yet
- Article Abou Shanab2011Document7 pagesArticle Abou Shanab2011Benelux ModeNo ratings yet
- International Biodeterioration & Biodegradation: Toruloides Using An Open-Bubble-Column Reactor Under Non-AsepticDocument10 pagesInternational Biodeterioration & Biodegradation: Toruloides Using An Open-Bubble-Column Reactor Under Non-AsepticFalcon ChauNo ratings yet
- Fabrication and Characterization of Poly Lactic Acid (PLA) - Starch Based Bioplastic CompositesDocument11 pagesFabrication and Characterization of Poly Lactic Acid (PLA) - Starch Based Bioplastic CompositesAndrea VONo ratings yet
- Fei 2016Document8 pagesFei 2016Davide Di ZioNo ratings yet
- FTB 54 489Document8 pagesFTB 54 489Nurjihan FaNo ratings yet
- Application of Green ChemistryDocument19 pagesApplication of Green ChemistryKumaran RNo ratings yet
- DownloadDocument12 pagesDownloadjhoanna grace nideaNo ratings yet
- Isolation of Poly-β-hydroxybutyrate (PHB) Producing Bacteria from Sago and Sugarcane Solid WasteDocument10 pagesIsolation of Poly-β-hydroxybutyrate (PHB) Producing Bacteria from Sago and Sugarcane Solid WasteFiqriansyahwahabNo ratings yet
- Biological Diversity in Aerated Facultative LagoonDocument17 pagesBiological Diversity in Aerated Facultative LagoonJaderson CarvalhoNo ratings yet
- Bioresource Technology: Contents Lists Available atDocument7 pagesBioresource Technology: Contents Lists Available atAtchaya அட்சயாNo ratings yet
- Kanjan A Chump Ol 2013Document12 pagesKanjan A Chump Ol 2013Kaomi CarbajalNo ratings yet
- Rayhane Et Al., 2018. From Flasks To Single Used Bioreactor Scale-Up of Solid State FermentationDocument9 pagesRayhane Et Al., 2018. From Flasks To Single Used Bioreactor Scale-Up of Solid State FermentationArely PradoNo ratings yet
- 10 1002@app 50412Document10 pages10 1002@app 50412Tuyul SatuNo ratings yet
- Production of Poly (3-Hydroxybutyrate) From A Dairy Industry Wastewater Using Bacillus Subtilis EPAH18: Bioprocess Development and SimulationDocument9 pagesProduction of Poly (3-Hydroxybutyrate) From A Dairy Industry Wastewater Using Bacillus Subtilis EPAH18: Bioprocess Development and Simulationroseta0218No ratings yet
- 1 s2.0 S0926669023002674 MainDocument11 pages1 s2.0 S0926669023002674 MainOmar KHELILNo ratings yet
- Bioresource Technology: Malik S.Y. Haddadin, Jamal Haddadin, Omar I. Arabiyat, Butros HattarDocument10 pagesBioresource Technology: Malik S.Y. Haddadin, Jamal Haddadin, Omar I. Arabiyat, Butros HattarRodrigo TorresNo ratings yet
- Bhuwal 2013Document10 pagesBhuwal 2013keysofiNo ratings yet
- Production of The Lactic Acid From Mango Peel Waste - Factorial ExperimentDocument7 pagesProduction of The Lactic Acid From Mango Peel Waste - Factorial ExperimentVj ReddyNo ratings yet
- International Journal of Engineering and Science Invention (IJESI)Document5 pagesInternational Journal of Engineering and Science Invention (IJESI)inventionjournalsNo ratings yet
- Development of Biomass-Based Plastic Resins From Natural Algal PolysaccharidesDocument72 pagesDevelopment of Biomass-Based Plastic Resins From Natural Algal Polysaccharidesjustin_domingo_2100% (2)
- Chemrj 2016 01 04 01 10Document10 pagesChemrj 2016 01 04 01 10editor chemrjNo ratings yet
- 14.Applied-BioPlastic A Better-Mukti GilDocument6 pages14.Applied-BioPlastic A Better-Mukti GilImpact JournalsNo ratings yet
- Biogas Production From Food Waste ThesisDocument5 pagesBiogas Production From Food Waste Thesiskualxkiig100% (1)
- Research Article Bacillus SubtilisDocument10 pagesResearch Article Bacillus SubtilisDương Nguyễn Thùy DungNo ratings yet
- 2018) Finding of Novel Lactate Utilizing Bacillus Sp. YHY22 and Its Evaluation For Polyhydroxybutyrate (PHB) ProductionDocument9 pages2018) Finding of Novel Lactate Utilizing Bacillus Sp. YHY22 and Its Evaluation For Polyhydroxybutyrate (PHB) Productionadmos0319No ratings yet
- 10 11648 J Ajpst 20200602 12Document7 pages10 11648 J Ajpst 20200602 12data.caindog.swuNo ratings yet
- Jurnal 5-Bio Waste KitosanDocument12 pagesJurnal 5-Bio Waste KitosanNurhaliza YanuartiNo ratings yet
- Production of Ethanol by Fermentation Process by Using YeastDocument9 pagesProduction of Ethanol by Fermentation Process by Using Yeastdeepu kumarNo ratings yet
- Bioethanols 2Document39 pagesBioethanols 2ahmad buysyNo ratings yet
- Fungi and Lignocellulosic BiomassFrom EverandFungi and Lignocellulosic BiomassChristian P KubicekNo ratings yet
- ZMQ 9 H 2 CM5 I0 CDocument666 pagesZMQ 9 H 2 CM5 I0 Cashuna249396No ratings yet
- Review of Xanthan Gum Production From Unmodified Starches-2006Document11 pagesReview of Xanthan Gum Production From Unmodified Starches-2006ashuna249396No ratings yet
- Xanthan Gum ScifinderDocument15 pagesXanthan Gum Scifinderashuna249396No ratings yet
- BT 0306Document3 pagesBT 0306ashuna249396No ratings yet
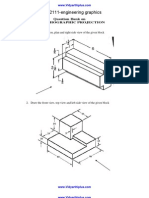
- GE2111-engineering Graphics: 1. Draw The Elevation, Plan and Right Side View of The Given BlockDocument4 pagesGE2111-engineering Graphics: 1. Draw The Elevation, Plan and Right Side View of The Given Blockashuna249396No ratings yet
- Production of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-4-Hydroxybutyrate) by Ralstonia Eutropha From Soybean OilDocument6 pagesProduction of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-4-Hydroxybutyrate) by Ralstonia Eutropha From Soybean Oilashuna249396No ratings yet
- Model-Based Fed-Batch Cultivation of R. Eutropha For Enhanced Biopolymer ProductionDocument8 pagesModel-Based Fed-Batch Cultivation of R. Eutropha For Enhanced Biopolymer Productionashuna249396No ratings yet
- Appl. Environ. Microbiol. 2009 Ibrahim 6222 31Document10 pagesAppl. Environ. Microbiol. 2009 Ibrahim 6222 31ashuna249396No ratings yet
- Cobas 4800 For HPV Detection and GenotypingDocument2 pagesCobas 4800 For HPV Detection and GenotypingMolecular_Diagnostics_KKUH0% (1)
- The Sauropods Evolution and Paleobiology - Kristina Curry Rogers Amp Amp Jeffrey Wilson PDFDocument360 pagesThe Sauropods Evolution and Paleobiology - Kristina Curry Rogers Amp Amp Jeffrey Wilson PDFManu Adri100% (4)
- Immunosero Notes 2Document20 pagesImmunosero Notes 2Rakim MikarNo ratings yet
- Ecas Clima Report 2018Document24 pagesEcas Clima Report 2018Aishwarya MathurNo ratings yet
- General Biology - Cell TheoryDocument4 pagesGeneral Biology - Cell TheorySage Of WholesomenessNo ratings yet
- Lesson 2 Upper ExtremitiesDocument23 pagesLesson 2 Upper ExtremitiesMARY JANE ANGELICA SEVANo ratings yet
- Bacteria ClassificationDocument395 pagesBacteria ClassificationJitendra MishraNo ratings yet
- SGBAU B.Pharm 1 SEM Human-Anatomy-n-Physiology-I 2018Document2 pagesSGBAU B.Pharm 1 SEM Human-Anatomy-n-Physiology-I 2018Abhay DeulkarNo ratings yet
- STS Project 2Document27 pagesSTS Project 2Diana CalderonNo ratings yet
- 12 White Blood Cell Counting Techniques in BirdsDocument5 pages12 White Blood Cell Counting Techniques in BirdsPablo LópezNo ratings yet
- Personal StatementDocument2 pagesPersonal StatementDaniela PerezNo ratings yet
- NSB Bulletin Feb. 2014Document153 pagesNSB Bulletin Feb. 2014nrpindiaNo ratings yet
- DuPont Distillase SSF Web enDocument2 pagesDuPont Distillase SSF Web ensushantNo ratings yet
- Soal Bahasa InggrisDocument5 pagesSoal Bahasa InggrisBintang MareetaNo ratings yet
- Jipmer DissertationsDocument8 pagesJipmer DissertationsPayToDoMyPaperUK100% (1)
- Pathologic Basis of Lumbar Radicular Pain: BackgroundDocument8 pagesPathologic Basis of Lumbar Radicular Pain: BackgroundVizaNo ratings yet
- Detection of Feline Panleukopenia With Antigen Test KitDocument2 pagesDetection of Feline Panleukopenia With Antigen Test KitMuhammad ihwanul usliminNo ratings yet
- Georges Cuvier's Theory of Correlation of PartsDocument5 pagesGeorges Cuvier's Theory of Correlation of PartsSoc SaballaNo ratings yet
- SUBHADIPA MAJUMDER2022-07-22Cell PotencyDocument2 pagesSUBHADIPA MAJUMDER2022-07-22Cell PotencySuvNo ratings yet
- Jacqueline Badgery Resume 2Document1 pageJacqueline Badgery Resume 2api-445750548No ratings yet
- Type 3 - Summary CompletionDocument7 pagesType 3 - Summary CompletionAnh ThuNo ratings yet
- 2021 Expt 7 Pre Lab - Diversity of BacteriacDocument2 pages2021 Expt 7 Pre Lab - Diversity of BacteriacjijiqNo ratings yet
- FlowerChart PDFDocument7 pagesFlowerChart PDFarthyNo ratings yet
- Orientation To The Human Body SlidesDocument24 pagesOrientation To The Human Body SlidesSamantha Kaye ReclaNo ratings yet
- Root Apex and It's SignificanceDocument33 pagesRoot Apex and It's Significancecitidotnet95% (20)
- 4life Transfer Factor TrifactorDocument2 pages4life Transfer Factor TrifactorYemsar AlhadarNo ratings yet
- Ether Lipids in Biomembranes: Fritz PaltaufDocument39 pagesEther Lipids in Biomembranes: Fritz PaltaufdnatechNo ratings yet
- Oig Method Validation Procedure 01 PDFDocument10 pagesOig Method Validation Procedure 01 PDFMargaretaSianneNo ratings yet
- Fascia and Primo Vascular SystemDocument7 pagesFascia and Primo Vascular SystemVladimir Rodrigues de LimaNo ratings yet