Download as pdf or txt
You might also like
- Module 3 - Fire SafetyDocument5 pagesModule 3 - Fire SafetyvonsonNo ratings yet
- 35 245-252 333-13 3910 Waldo Sven Lavado Casimiro CorrDocument8 pages35 245-252 333-13 3910 Waldo Sven Lavado Casimiro CorrStela CarreraNo ratings yet
- DroughtDocument22 pagesDroughtfangoursolution10No ratings yet
- TerraClimate, A High-Resolution Global Dataset of Monthly Climate and Climatic Water Balance From 1958-2015Document12 pagesTerraClimate, A High-Resolution Global Dataset of Monthly Climate and Climatic Water Balance From 1958-2015Bianca FazacasNo ratings yet
- (15257541 - Journal of Hydrometeorology) The Version-2 Global Precipitation Climatology Project (GPCP) Monthly Precipitation Analysis (1979-Present)Document21 pages(15257541 - Journal of Hydrometeorology) The Version-2 Global Precipitation Climatology Project (GPCP) Monthly Precipitation Analysis (1979-Present)lbarretoNo ratings yet
- The Relationship Between NDVI and Precipitation On The Tibetan PlateauDocument10 pagesThe Relationship Between NDVI and Precipitation On The Tibetan PlateauBetNo ratings yet
- Lazcarro 2020 IOP Conf. Ser. Earth Environ. Sci. 451 012023Document10 pagesLazcarro 2020 IOP Conf. Ser. Earth Environ. Sci. 451 012023Valentina DavydovaNo ratings yet
- Intl Journal of Climatology - 2022 - Xavier - New Improved Brazilian Daily Weather Gridded Data 1961 2020Document15 pagesIntl Journal of Climatology - 2022 - Xavier - New Improved Brazilian Daily Weather Gridded Data 1961 2020Carlos BenicioNo ratings yet
- Impact of Land Use and Climate Change On Surface Runoff Using GIS and SWAT Model - A Case StudyDocument10 pagesImpact of Land Use and Climate Change On Surface Runoff Using GIS and SWAT Model - A Case StudyAbhishëk ChhonkarNo ratings yet
- Spatial Modeling and Interpolation of Monthly Temperature Using KrigingDocument11 pagesSpatial Modeling and Interpolation of Monthly Temperature Using KrigingvisaslaveNo ratings yet
- EARSeL Symposium 2010 - 17 03 PDFDocument8 pagesEARSeL Symposium 2010 - 17 03 PDFjhongs1988No ratings yet
- Very High Resolution Interpolated Climate Surfaces For Global Land AreasDocument14 pagesVery High Resolution Interpolated Climate Surfaces For Global Land AreasRijhalVHirusNo ratings yet
- Observational Datasets Used in Climate Studies: Appendix ADocument6 pagesObservational Datasets Used in Climate Studies: Appendix ADinesh GaikwadNo ratings yet
- Observational Datasets Used in Climate Studies: Appendix ADocument6 pagesObservational Datasets Used in Climate Studies: Appendix AagapocorpNo ratings yet
- 1 s2.0 S0048969724031991 MainDocument12 pages1 s2.0 S0048969724031991 MainAdil SALHINo ratings yet
- RUSLEPerdidade Suelo SIGDocument13 pagesRUSLEPerdidade Suelo SIGlmartinezbNo ratings yet
- Hess 21 1295 2017Document26 pagesHess 21 1295 2017sandraNo ratings yet
- Travel TimeDocument17 pagesTravel TimeSudharsananPRSNo ratings yet
- Agriculture Journal Presentation of Four Centennial-Long Global Gridded Datasets of The Standardized Precipitation IndexDocument13 pagesAgriculture Journal Presentation of Four Centennial-Long Global Gridded Datasets of The Standardized Precipitation IndexAgriculture JournalNo ratings yet
- Shibu 2007Document2 pagesShibu 2007Freddy AngelitoNo ratings yet
- A New Methodology For Drought Vulnerability Assessment Using SPI (Standardized Precipitation Index)Document8 pagesA New Methodology For Drought Vulnerability Assessment Using SPI (Standardized Precipitation Index)Amin MojiriNo ratings yet
- Agudelo P.A. 2004 - Analysis of Spatial Distribution in Tropospheric Temperature TrendsDocument5 pagesAgudelo P.A. 2004 - Analysis of Spatial Distribution in Tropospheric Temperature TrendserikNo ratings yet
- 1984 WGEN RichardsonDocument88 pages1984 WGEN RichardsonAbdusalam IdirisNo ratings yet
- Benchmarking Optical/thermal Satellite Imagery For Estimating Evapotranspiration and Soil Moisture in Decision Support ToolsDocument31 pagesBenchmarking Optical/thermal Satellite Imagery For Estimating Evapotranspiration and Soil Moisture in Decision Support ToolsJanMHHendrickxNo ratings yet
- Gruber Et Al 2000 Bull GPCP PDFDocument14 pagesGruber Et Al 2000 Bull GPCP PDFvagoulNo ratings yet
- Usunoff 2Document9 pagesUsunoff 2SudharsananPRSNo ratings yet
- Development of Scale-Free Climate Data For Western Canada For Use in Resource ManagementDocument15 pagesDevelopment of Scale-Free Climate Data For Western Canada For Use in Resource Managementwilujeng nur hidayatNo ratings yet
- 14 Santillan0048 AuthorDocument12 pages14 Santillan0048 AuthorkamutmazNo ratings yet
- Areal Extent of Drought PDFDocument7 pagesAreal Extent of Drought PDFMinn KoKo KyawNo ratings yet
- Remote SensingDocument35 pagesRemote SensingantoniocamargoNo ratings yet
- M Onitoring Land-Cover Changes: A Com Parison of Change Detection TechniquesDocument14 pagesM Onitoring Land-Cover Changes: A Com Parison of Change Detection Techniquesikhsan07No ratings yet
- Li 2004Document14 pagesLi 2004Alfredo DezoNo ratings yet
- Estimation of Hydraulic Modeling Parameters of Poorly Gauged Basins Using Remote Sensing and GISDocument7 pagesEstimation of Hydraulic Modeling Parameters of Poorly Gauged Basins Using Remote Sensing and GISDeniz Ozde YamanNo ratings yet
- KC and NDVI 01Document15 pagesKC and NDVI 01Ranveer SinggNo ratings yet
- Papa David 2013Document14 pagesPapa David 2013Klissman Morales OlabarreraNo ratings yet
- Nie2018 Article AGlobalHydrologicalDroughtIndeDocument16 pagesNie2018 Article AGlobalHydrologicalDroughtIndeMurat BektaşoğluNo ratings yet
- Paper Analisa Bencana Di Banjir Kota PadangDocument8 pagesPaper Analisa Bencana Di Banjir Kota Padangrayen1212No ratings yet
- Evaluation of Remote Sensing-Based Irrigated Area Map For The Conterminous United StatesDocument13 pagesEvaluation of Remote Sensing-Based Irrigated Area Map For The Conterminous United StatesMinn Thu NaingNo ratings yet
- Spatial Distribution of Groundwater Quality With Geostatistics (Case Study: Yazd-Ardakan Plain)Document9 pagesSpatial Distribution of Groundwater Quality With Geostatistics (Case Study: Yazd-Ardakan Plain)Solomon P. GbanieNo ratings yet
- (J) Spaceborne GNSS - R Retrieving On Global Soil Moisture ApproachedDocument6 pages(J) Spaceborne GNSS - R Retrieving On Global Soil Moisture ApproachedAhmed WasiuNo ratings yet
- Remote Sensing of Environment: SciencedirectDocument13 pagesRemote Sensing of Environment: SciencedirectEdmundo CanchariNo ratings yet
- Rain4peDocument28 pagesRain4peCristianNo ratings yet
- Delgado2021 Chapter InterrelationsBetweenSoilErosiDocument12 pagesDelgado2021 Chapter InterrelationsBetweenSoilErosiDaniel DelgadoNo ratings yet
- Spatial Variability of Precipitation in SpainDocument7 pagesSpatial Variability of Precipitation in SpainZeNo ratings yet
- Vegetation Cover Annual Changes Based On Modis/Terra Ndvi in The Three Gorges Reservoir AreaDocument4 pagesVegetation Cover Annual Changes Based On Modis/Terra Ndvi in The Three Gorges Reservoir AreaAnyak2014No ratings yet
- Application of Vegetation Index and Brightness Temperature For Drought DetectionDocument10 pagesApplication of Vegetation Index and Brightness Temperature For Drought DetectionnasefgisNo ratings yet
- Flood Modelling PDFDocument12 pagesFlood Modelling PDFShashika IreshNo ratings yet
- (15200477 - Bulletin of The American Meteorological Society) The Global Precipitation Climatology Project (GPCP) Combined Precipitation DatasetDocument16 pages(15200477 - Bulletin of The American Meteorological Society) The Global Precipitation Climatology Project (GPCP) Combined Precipitation DatasetALANA RAQUEL PIRESNo ratings yet
- A New Topographic Index To Quantify DownDocument6 pagesA New Topographic Index To Quantify DownAdriano CHAVES DE FRANÇANo ratings yet
- Remotesensing 05 01425Document14 pagesRemotesensing 05 01425ceice2013No ratings yet
- AYBAR Et Al. 2019 - Construction High-Res Gridded Rainfall Dataset For Peru From 1981 To The Present DayDocument17 pagesAYBAR Et Al. 2019 - Construction High-Res Gridded Rainfall Dataset For Peru From 1981 To The Present DayFabian DrenkhanNo ratings yet
- Lab 6 ReportDocument7 pagesLab 6 Reportapi-275943676No ratings yet
- Simulating The Impact of Climate Change and Land Use On Groundwater Percolation RateDocument4 pagesSimulating The Impact of Climate Change and Land Use On Groundwater Percolation RateAJER JOURNALNo ratings yet
- PGBinder MAPDocument13 pagesPGBinder MAPCleber FaccinNo ratings yet
- Water 10 00213Document23 pagesWater 10 00213antoniocamargoNo ratings yet
- Validation Andes Sat RainDocument12 pagesValidation Andes Sat RainantoniocamargoNo ratings yet
- Vicente-Serrano Etal 2010Document18 pagesVicente-Serrano Etal 2010wacotc2163No ratings yet
- Testing Di Single Beam Echosounder River Surveying. Case Study: Siret RiverDocument21 pagesTesting Di Single Beam Echosounder River Surveying. Case Study: Siret RiverppallareNo ratings yet
- Hydraulic Modelling Using Hec-Ras and GiDocument5 pagesHydraulic Modelling Using Hec-Ras and GijgstormNo ratings yet
- Climate Finance as an Instrument to Promote the Green Growth in Developing CountriesFrom EverandClimate Finance as an Instrument to Promote the Green Growth in Developing CountriesNo ratings yet
- NIPCC vs. IPCC: Addressing the Disparity between Climate Models and Observations: Testing the Hypothesis of Anthropogenic Global WarmingFrom EverandNIPCC vs. IPCC: Addressing the Disparity between Climate Models and Observations: Testing the Hypothesis of Anthropogenic Global WarmingRating: 3 out of 5 stars3/5 (2)
- Guía Rápida de Las Aves de Los Tuxtlas: October 2015Document3 pagesGuía Rápida de Las Aves de Los Tuxtlas: October 2015rbyq9No ratings yet
- Biological Diversity: Unit 10Document29 pagesBiological Diversity: Unit 10rbyq9No ratings yet
- Saving Our Shared Birds: Partners in Flight Tri-National Vision For Landbird ConservationDocument52 pagesSaving Our Shared Birds: Partners in Flight Tri-National Vision For Landbird Conservationrbyq9No ratings yet
- BIO 1020 Unit 11Document23 pagesBIO 1020 Unit 11rbyq9No ratings yet
- Stable Isotope Analysis of Fall Migration Stopover by Six Passerine Species in An Inland Pitch Pine-Scrub Oak BarrenDocument10 pagesStable Isotope Analysis of Fall Migration Stopover by Six Passerine Species in An Inland Pitch Pine-Scrub Oak Barrenrbyq9No ratings yet
- Excubitorides) Wintering in MexicoDocument6 pagesExcubitorides) Wintering in Mexicorbyq9No ratings yet
- Unit 6 1Document21 pagesUnit 6 1rbyq90% (1)
- 0004-8038 (2006) 123 (822 Agatet) 2.0Document15 pages0004-8038 (2006) 123 (822 Agatet) 2.0rbyq9No ratings yet
- Assigning birds to geographic origin using feather hydrogen isotope ratios (δ H) : importance of year, age, and habitatDocument7 pagesAssigning birds to geographic origin using feather hydrogen isotope ratios (δ H) : importance of year, age, and habitatrbyq9No ratings yet
- Metabolism Exercises 2 and 3Document1 pageMetabolism Exercises 2 and 3rbyq9No ratings yet
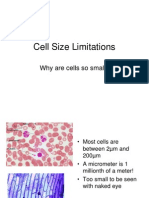
- Cell Size Limitations: Why Are Cells So Small?Document9 pagesCell Size Limitations: Why Are Cells So Small?rbyq9No ratings yet
- General Biology Labmake Up Worksheet Unit 8Document1 pageGeneral Biology Labmake Up Worksheet Unit 8rbyq9No ratings yet
- General Biology Lab Makeup Worksheet Unit 1 PDFDocument1 pageGeneral Biology Lab Makeup Worksheet Unit 1 PDFrbyq9No ratings yet
- Cell Structure and FunctionDocument9 pagesCell Structure and Functionrbyq9No ratings yet
- Lab03 OsmosisDocument7 pagesLab03 Osmosisrbyq90% (2)
- Hiradc Om - Talian AtasDocument57 pagesHiradc Om - Talian AtasapnadiNo ratings yet
- OS6900 AOS 8.9.107.R02 Specifications GuideDocument102 pagesOS6900 AOS 8.9.107.R02 Specifications Guidemenage.gillesNo ratings yet
- Art Collector I105 07.09 2023Document236 pagesArt Collector I105 07.09 2023JjxpresidentNo ratings yet
- Home Cmplab HTML Bbs Data Pds0085 1Document9 pagesHome Cmplab HTML Bbs Data Pds0085 1呂昕儒No ratings yet
- PLKDocument13 pagesPLKtunggal KecerNo ratings yet
- A Survey of Field Applications of Gel Treatments For Water ShutoffDocument11 pagesA Survey of Field Applications of Gel Treatments For Water ShutoffBình TrầnNo ratings yet
- Voltage Source Converter Based HVDC: Transmission and DistributionDocument4 pagesVoltage Source Converter Based HVDC: Transmission and DistributionAshok kumar RNo ratings yet
- Cromatografi e TextDocument8 pagesCromatografi e TextRaunciucGianinaNo ratings yet
- Wintergreen Phys-Ed 2012Document226 pagesWintergreen Phys-Ed 2012natasha_bretonNo ratings yet
- DB Reg SK GBDocument2 pagesDB Reg SK GBABDELLAHABDELLAH2010No ratings yet
- 1979 Herb Fitch - Beyond Time Seminar 4 - Soul CenteringDocument16 pages1979 Herb Fitch - Beyond Time Seminar 4 - Soul CenteringwilliamNo ratings yet
- MSFS Airbus Paintkit ManualDocument25 pagesMSFS Airbus Paintkit ManualNguyen Duc BinhNo ratings yet
- Mengistu 2016Document13 pagesMengistu 2016Dimas WahyuNo ratings yet
- SurinameDocument9 pagesSurinameReza Pall MallNo ratings yet
- (SOSC1960) (2012) (S) Quiz - jqj28 97890Document3 pages(SOSC1960) (2012) (S) Quiz - jqj28 97890alex61937No ratings yet
- 2021 Sustainability ReportDocument101 pages2021 Sustainability ReportGrupo DOCStore Brasil LtdaNo ratings yet
- Exp#1 Diode Characteristics V2Document12 pagesExp#1 Diode Characteristics V2Shehda ZahdaNo ratings yet
- Mark Scheme (Results) January 2007: GCE Biology (8040/9040)Document12 pagesMark Scheme (Results) January 2007: GCE Biology (8040/9040)Fathmath MohamedNo ratings yet
- Lichfield Sutton Coldfield Birmingham Long Bridge RedditchDocument4 pagesLichfield Sutton Coldfield Birmingham Long Bridge RedditchLouisMichaelNo ratings yet
- Uses of Nanotechnology in Environmental Technology in HessenDocument60 pagesUses of Nanotechnology in Environmental Technology in Hessenfelipe de jesus juarez torresNo ratings yet
- Exercise CONTEXT CLUESDocument2 pagesExercise CONTEXT CLUESSri MulyaniNo ratings yet
- Design Principles For Thick CylindersDocument18 pagesDesign Principles For Thick CylindersDinesh VaghelaNo ratings yet
- Joseph W. Mcmoneagle: Interview #001Document8 pagesJoseph W. Mcmoneagle: Interview #001n3ncomNo ratings yet
- Cooling Load Calculation Through Transmission and ProductDocument4 pagesCooling Load Calculation Through Transmission and ProductKenneth CariñoNo ratings yet
- Assignment - Condenser and Nozzle-1Document2 pagesAssignment - Condenser and Nozzle-1Harshil ChaddhaNo ratings yet
- Medical Oxygen Generator FlyerDocument2 pagesMedical Oxygen Generator FlyertahatekriNo ratings yet
- Class 11 Cell Cycle and Cell DivisionDocument2 pagesClass 11 Cell Cycle and Cell Divisionjeeneet.alwalNo ratings yet
- Real Life - The Body and Related VerbsDocument2 pagesReal Life - The Body and Related VerbsSuper JudyNo ratings yet
- Importance of Lohri FestivalDocument7 pagesImportance of Lohri FestivalThe United IndianNo ratings yet