Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Miller - The Drama of The Gifted ChildDocument3 pagesMiller - The Drama of The Gifted ChildTopan Setiadi83% (6)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Body DressingDocument263 pagesBody Dressingvijaykumarmv100% (4)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Prelim Examination: Select OneDocument4 pagesPrelim Examination: Select OneAxel Cabornay100% (3)
- Classic D&D Races For D&D 5th EditionDocument7 pagesClassic D&D Races For D&D 5th EditionRyan Lowder100% (11)
- Public Relation in HospitalDocument6 pagesPublic Relation in HospitalAnusha Verghese80% (5)
- The Trip of The Tip: Understanding The Growth Cone MachineryDocument12 pagesThe Trip of The Tip: Understanding The Growth Cone MachinerycstaiiNo ratings yet
- Science:, 1959 (2002) Barry J. DicksonDocument8 pagesScience:, 1959 (2002) Barry J. DicksoncstaiiNo ratings yet
- Mechanics of Brain TissueDocument13 pagesMechanics of Brain TissuecstaiiNo ratings yet
- Self-Organization of Myosin II in Reconstituted Actomyosin BundlesDocument10 pagesSelf-Organization of Myosin II in Reconstituted Actomyosin BundlescstaiiNo ratings yet
- Balduin I 2017Document11 pagesBalduin I 2017cstaiiNo ratings yet
- MTF270-Turbulence Modeling-Large Eddy Simulations PDFDocument29 pagesMTF270-Turbulence Modeling-Large Eddy Simulations PDFkulov1592No ratings yet
- Guadines v. SandiganbayanDocument13 pagesGuadines v. SandiganbayanDenise CuizonNo ratings yet
- New Rich Text DocumentDocument4 pagesNew Rich Text Documentprajjwal singhNo ratings yet
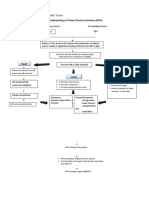
- Pathophysiology of Patent Ductus Arteroisus (PDA)Document2 pagesPathophysiology of Patent Ductus Arteroisus (PDA)Rodel Yacas100% (1)
- Self-Assessment 9.8: For Each Learning Objective, Tick ( ) The Box That Best Matches Your AbilityDocument1 pageSelf-Assessment 9.8: For Each Learning Objective, Tick ( ) The Box That Best Matches Your AbilityКатерина КравченкоNo ratings yet
- W Iser - Towards Literary AnthropologyDocument5 pagesW Iser - Towards Literary AnthropologyMaria Teresa De PalmaNo ratings yet
- Complaint - Spence v. Insomnia CookiesDocument13 pagesComplaint - Spence v. Insomnia CookiespospislawNo ratings yet
- Comparison Between Task Oriented Training and Proprioceptiveneuromuscular Facilitation Exercises On Lower Extremity Function Ince 2165 7025 1000291Document9 pagesComparison Between Task Oriented Training and Proprioceptiveneuromuscular Facilitation Exercises On Lower Extremity Function Ince 2165 7025 1000291Gopi KrishnanNo ratings yet
- Research ProposalDocument9 pagesResearch Proposalaarya chandrakar100% (1)
- Crim Law II ReviewerDocument157 pagesCrim Law II ReviewerJierah ManahanNo ratings yet
- The Changing Marketing LandscapeDocument14 pagesThe Changing Marketing Landscapeagatha linusNo ratings yet
- Dyck CommentaryDocument32 pagesDyck CommentaryRichardNo ratings yet
- 4 Huge Benefits of Meditation (And How To Get Started Today)Document16 pages4 Huge Benefits of Meditation (And How To Get Started Today)Jia Huay Chang (MY)No ratings yet
- Seminar Progressive Patient CareDocument16 pagesSeminar Progressive Patient CareGayathri R100% (1)
- The Iron Man (CH 2 Part 1) - Word DetectiveDocument5 pagesThe Iron Man (CH 2 Part 1) - Word DetectiveNur Izyani RasdiNo ratings yet
- My First Writing - Workbook 2 - Answer Key (E-Future)Document26 pagesMy First Writing - Workbook 2 - Answer Key (E-Future)Mona MohamedelsayedNo ratings yet
- Aaron Beck's Cognitive DevelopmentDocument7 pagesAaron Beck's Cognitive DevelopmentPriscilla KiemoNo ratings yet
- 7 Major Elements of Communication ProcessDocument2 pages7 Major Elements of Communication ProcessEarl John Fajarito Rafael100% (3)
- Comparison of Available Shear Strength Models For Non ConformingDocument16 pagesComparison of Available Shear Strength Models For Non ConformingSaad KhanNo ratings yet
- CivilsyllDocument131 pagesCivilsyllYashwanth YashuNo ratings yet
- PREVIEW Cognos SDK Guide by BI CentreDocument12 pagesPREVIEW Cognos SDK Guide by BI CentreHadi AamirNo ratings yet
- Presentation On "Public Key Cryptography": Submitted ByDocument30 pagesPresentation On "Public Key Cryptography": Submitted Bykritagya chandraNo ratings yet
- Joshua John Atkinson Sentencing NotesDocument2 pagesJoshua John Atkinson Sentencing NotesexaminertasNo ratings yet
- (1994) The Stationary Bootstrap - Politis and RomanoDocument12 pages(1994) The Stationary Bootstrap - Politis and RomanoGabe LinNo ratings yet
- The Vietnam War: Pre-ReadingDocument8 pagesThe Vietnam War: Pre-ReadingLinh Thuy NguyenNo ratings yet