Download as pdf or txt
You might also like
- Ganong Textbook of Medical PhysiologyDocument98 pagesGanong Textbook of Medical PhysiologyJanie-Vi Gorospe100% (1)
- NCP HemothoraxDocument3 pagesNCP HemothoraxMichael John F. NatividadNo ratings yet
- Review Tubulin-Dynein System in Agellar and Ciliary MovementDocument19 pagesReview Tubulin-Dynein System in Agellar and Ciliary Movementsama EmadNo ratings yet
- Structure of Microtubules Amd ActinsDocument19 pagesStructure of Microtubules Amd Actinspreetakaran12No ratings yet
- Myosin Motors Drive Long Range AlignmentDocument12 pagesMyosin Motors Drive Long Range AlignmentAlexNo ratings yet
- ChemBioChem - 2002 - MC Henry - A Non Antibacterial Oxazolidinone Derivative That Inhibits Epithelial Cell Sheet MigrationDocument7 pagesChemBioChem - 2002 - MC Henry - A Non Antibacterial Oxazolidinone Derivative That Inhibits Epithelial Cell Sheet MigrationGabriel FenteanyNo ratings yet
- Acetylated Microtubules Are Preferentially Bundled Leading - 2017 - BiophysicalDocument10 pagesAcetylated Microtubules Are Preferentially Bundled Leading - 2017 - BiophysicalDana MateiNo ratings yet
- 03 Tabas I (2002) Cholesterol in Health and DiseaseDocument8 pages03 Tabas I (2002) Cholesterol in Health and DiseaseCristobal Aguila RebolledoNo ratings yet
- Hirano BodiesDocument10 pagesHirano BodiesAlvaro Martin Bonsak ArceNo ratings yet
- Err 265Document9 pagesErr 265Najmi HanifahNo ratings yet
- Traffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlDocument8 pagesTraffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlPinaki NayakNo ratings yet
- Microfilament: in Vitro Self-AssemblyDocument7 pagesMicrofilament: in Vitro Self-AssemblyABHISHEK RATHODNo ratings yet
- Cell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Document26 pagesCell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Rajni KumariNo ratings yet
- Self-Organization of Myosin II in Reconstituted Actomyosin BundlesDocument10 pagesSelf-Organization of Myosin II in Reconstituted Actomyosin BundlescstaiiNo ratings yet
- Lecture 3 - Chapter 8-Cytoskeleton ADocument75 pagesLecture 3 - Chapter 8-Cytoskeleton AKw Chan33% (3)
- Pys PDFDocument5 pagesPys PDFMD shah KhanNo ratings yet
- 1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesDocument19 pages1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesMD shah KhanNo ratings yet
- The International Journal of Biochemistry & Cell Biology: TropoelastinDocument4 pagesThe International Journal of Biochemistry & Cell Biology: TropoelastinNg SoohsienNo ratings yet
- Chapter 16 - The Cytoskeleton - 112612Document56 pagesChapter 16 - The Cytoskeleton - 112612Sohan rautNo ratings yet
- Cholesterol in Health and DiseaseDocument8 pagesCholesterol in Health and DiseaseErwin K Sjah AlamNo ratings yet
- Reflection LoueDocument11 pagesReflection LoueDaisy C. LabradaNo ratings yet
- Spermatogénèse 2017Document11 pagesSpermatogénèse 2017amalaoumrani15No ratings yet
- 04 15 Motor Proteins MilliganDocument8 pages04 15 Motor Proteins Milliganعلى الله متوكلNo ratings yet
- Clathrin Is Involved in Organization of Mitotic Spindle and Phragmoplast As Well As in Endocytosis in Tobacco Cell CulturesDocument11 pagesClathrin Is Involved in Organization of Mitotic Spindle and Phragmoplast As Well As in Endocytosis in Tobacco Cell CulturesNuke Nur HidayatiNo ratings yet
- Vol 2 Iss 2 Page 5-13 Molecular Motors A Review ArticleDocument9 pagesVol 2 Iss 2 Page 5-13 Molecular Motors A Review ArticleEditorsNo ratings yet
- NIH Public Access: Author ManuscriptDocument18 pagesNIH Public Access: Author ManuscriptMaria IordacheNo ratings yet
- Aamir Nawaz KhanDocument20 pagesAamir Nawaz KhanBslssbxjxnbbxNo ratings yet
- Bio KimDocument6 pagesBio KimKarina SaputraNo ratings yet
- Pectin-Coated Titanium Implants Are Well-Tolerated in Vivo: 2009 Wiley Periodicals, IncDocument6 pagesPectin-Coated Titanium Implants Are Well-Tolerated in Vivo: 2009 Wiley Periodicals, Incbiomedicalengineer 27No ratings yet
- Topic: Cytokinesis: Etymology and PronunciationDocument6 pagesTopic: Cytokinesis: Etymology and PronunciationVindhya ShankerNo ratings yet
- 14 The CytoskeletonDocument7 pages14 The Cytoskeletonsatriamanullang880No ratings yet
- 10.1007@978 1 4614 3573 010Document28 pages10.1007@978 1 4614 3573 010Fiy Jannatin AliyahNo ratings yet
- Cilia and FlagellaDocument25 pagesCilia and FlagellaLia Savitri RomdaniNo ratings yet
- Chapter 2: Cell Structure and Cell Organisation: Textbook Focus Practise 2.1 (Page 22)Document8 pagesChapter 2: Cell Structure and Cell Organisation: Textbook Focus Practise 2.1 (Page 22)Siva RajahNo ratings yet
- Biology Paper 3Document12 pagesBiology Paper 3Fitri RamadhaniNo ratings yet
- Plant Cell Structure and Importance of Cell Organelles I Post Harvest PhysiologyDocument10 pagesPlant Cell Structure and Importance of Cell Organelles I Post Harvest Physiologychristom karungariNo ratings yet
- Aps 2005114Document11 pagesAps 2005114sosemir589No ratings yet
- Study GuideDocument64 pagesStudy GuideCatherine GrivotNo ratings yet
- He Cell 1Document34 pagesHe Cell 1Le Phuong LyNo ratings yet
- Physicochemical Characterization of Chitosan Nanoparticles: Electrokinetic and Stability BehaviorDocument8 pagesPhysicochemical Characterization of Chitosan Nanoparticles: Electrokinetic and Stability BehaviorMihaela HolbanNo ratings yet
- Inigo.j 01109151013Document27 pagesInigo.j 01109151013inigoNo ratings yet
- Plant Hormones. Methods and Protocols, 2nd EdnDocument1 pagePlant Hormones. Methods and Protocols, 2nd EdnThảo Quỳnh Anh HồNo ratings yet
- Structure and Function of Photosystem SupercomplexesDocument7 pagesStructure and Function of Photosystem Supercomplexessri dinaNo ratings yet
- Prokaryotic Cell, Cell That Lack A Nucleus and Membrane Bound Organelles. Myosin Is Responsible For ADocument2 pagesProkaryotic Cell, Cell That Lack A Nucleus and Membrane Bound Organelles. Myosin Is Responsible For AHàyyåt ChuadhryNo ratings yet
- Jesse Science DocumentDocument7 pagesJesse Science Documentapi-260887014No ratings yet
- Relating Structure To Mechanism in Creatine KinaseDocument21 pagesRelating Structure To Mechanism in Creatine KinaseAnusorn S NiiNo ratings yet
- Symmetry-Breaking of Animal CytokinesisDocument10 pagesSymmetry-Breaking of Animal CytokinesisLizbeth EspinosaNo ratings yet
- UNIT 2 (Unfinished)Document22 pagesUNIT 2 (Unfinished)青木ケイNo ratings yet
- CHAP 2. CSH TẾ BÀO - 232Document100 pagesCHAP 2. CSH TẾ BÀO - 232Nguyễn Anh QuangNo ratings yet
- Articulo HFMDocument38 pagesArticulo HFMLiliana MesaNo ratings yet
- bk978 0 7503 1302 5ch1Document31 pagesbk978 0 7503 1302 5ch1Vaibhav RAJ KUMAR GUPTANo ratings yet
- Signor Methods PaperDocument9 pagesSignor Methods PaperPrasetyaNo ratings yet
- Actomyosin Networks and Tissue MorphogenesisDocument5 pagesActomyosin Networks and Tissue MorphogenesisArkayan LahaNo ratings yet
- 2016 JhaP CPMoB 2016Document15 pages2016 JhaP CPMoB 2016Maria SunflowerNo ratings yet
- Mitochondrial Respiration and Redox Coupling in Articular ChondrocytesDocument14 pagesMitochondrial Respiration and Redox Coupling in Articular ChondrocytesndsjNo ratings yet
- Cytology CytoskeletonDocument48 pagesCytology CytoskeletonMitzel Sapalo0% (1)
- 7.3 Programmed Proteolysis: 7.3.1 General FeaturesDocument23 pages7.3 Programmed Proteolysis: 7.3.1 General FeaturesdragoncytoNo ratings yet
- Cytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDocument11 pagesCytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDario DiosNo ratings yet
- TMP 94 C1Document121 pagesTMP 94 C1FrontiersNo ratings yet
- Myosin, Actin & Contraction NotesDocument4 pagesMyosin, Actin & Contraction NotesGhazali BalochNo ratings yet
- Emerging Functions of Actins and Actin Binding Proteins in TrypanosomatidsDocument23 pagesEmerging Functions of Actins and Actin Binding Proteins in TrypanosomatidsRudi haris munandarNo ratings yet
- 1004 Belize BromeliaceaeDocument6 pages1004 Belize BromeliaceaeHarry JonitzNo ratings yet
- Traveset 2015 Bird-Flower Visitation Networks in The Galapagos PDFDocument6 pagesTraveset 2015 Bird-Flower Visitation Networks in The Galapagos PDFHarry JonitzNo ratings yet
- JackfruitDocument9 pagesJackfruitHarry JonitzNo ratings yet
- Ammonites EcuadorDocument10 pagesAmmonites EcuadorHarry JonitzNo ratings yet
- Menzel - The Role of Cytoskeleton On PolarityDocument5 pagesMenzel - The Role of Cytoskeleton On PolarityHarry JonitzNo ratings yet
- DasycladalesDocument373 pagesDasycladalesHarry JonitzNo ratings yet
- Acuña 2013 First Record of Three Shark SpeciesDocument6 pagesAcuña 2013 First Record of Three Shark SpeciesHarry JonitzNo ratings yet
- System 1 Binaural Beats 0001Document12 pagesSystem 1 Binaural Beats 0001Golden SunriseNo ratings yet
- Animal BehaviourDocument5 pagesAnimal BehaviourthinaNo ratings yet
- Final Collected Questions of Gis 018Document10 pagesFinal Collected Questions of Gis 018Mohammad DarkhabaniNo ratings yet
- Orthodontics Ii (Lecture Note)Document29 pagesOrthodontics Ii (Lecture Note)Rufai HarunaNo ratings yet
- Grade 6: The Heart and Circulatory SystemDocument18 pagesGrade 6: The Heart and Circulatory SystemEstanislao Villanueva ElemNo ratings yet
- Definitive Airway Management After Prehospital Supraglottic Rescue Airway in Pediatric Trauma PDFDocument5 pagesDefinitive Airway Management After Prehospital Supraglottic Rescue Airway in Pediatric Trauma PDFikm fkunissulaNo ratings yet
- Biology SkeletonDocument36 pagesBiology SkeletonHariPriya JothimeenakshiNo ratings yet
- WORKSHOP No 3Document20 pagesWORKSHOP No 3Yainel RomeroNo ratings yet
- Human Reproduction by Garima Mam)Document40 pagesHuman Reproduction by Garima Mam)MOHD zAIDNo ratings yet
- Drug Study NCP SoapieDocument15 pagesDrug Study NCP Soapiemikrobyo_ng_wmsuNo ratings yet
- H&N Wk1 Face MusclesDocument10 pagesH&N Wk1 Face MusclesUloko ChristopherNo ratings yet
- Unit 10 Comfort Pain Rest and SleepDocument2 pagesUnit 10 Comfort Pain Rest and SleepAizan Syahirah NoordinNo ratings yet
- CME Brochure 21 Feb 2015Document2 pagesCME Brochure 21 Feb 2015ParameshParamamNo ratings yet
- Mold ToxinsDocument7 pagesMold ToxinsMikiatlantul100% (1)
- Bee Stings Immunology Allergy and Treatment Marterre PDFDocument9 pagesBee Stings Immunology Allergy and Treatment Marterre PDFOktaviana Sari DewiNo ratings yet
- Acneiform Disorders: Chapter 78:: Acne Vulgaris:: Carolyn Goh, Carol Cheng, George AgakDocument28 pagesAcneiform Disorders: Chapter 78:: Acne Vulgaris:: Carolyn Goh, Carol Cheng, George AgakTrustia RizqandaruNo ratings yet
- Acit TrainingDocument23 pagesAcit TrainingTheoNo ratings yet
- Arterial Blood Gas AnalysisDocument24 pagesArterial Blood Gas AnalysisDeepika PatelNo ratings yet
- Biotensegrity ReviewDocument19 pagesBiotensegrity ReviewTensegrity Wiki100% (3)
- Haematology PhysiologyDocument15 pagesHaematology PhysiologyOlivia LimNo ratings yet
- Hemorrhage NCPDocument4 pagesHemorrhage NCPElishaNo ratings yet
- Necrosis and Types of NecrosisDocument2 pagesNecrosis and Types of NecrosisHisham NesemNo ratings yet
- Bio 93 Midterm Review For The FinalDocument1 pageBio 93 Midterm Review For The Finalsarah_choi_21No ratings yet
- 39.2 Laparoscopic Adrenalectomy: Ahmad Assalia, M.D. Michel Gagner, M.D., FACS, FRCSCDocument13 pages39.2 Laparoscopic Adrenalectomy: Ahmad Assalia, M.D. Michel Gagner, M.D., FACS, FRCSCimherestudyingNo ratings yet
- The Spiral Engine of Locomotion: Five Primary Kinetic ChainsDocument12 pagesThe Spiral Engine of Locomotion: Five Primary Kinetic ChainsAbhishek VermaNo ratings yet
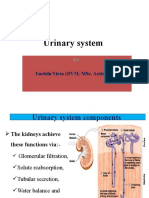
- Urinary System: Tsedalu Yirsa (DVM, MSC, Assist Prof.)Document18 pagesUrinary System: Tsedalu Yirsa (DVM, MSC, Assist Prof.)Mohammed OumerNo ratings yet
- Neuronal Integration and CurcuitsDocument5 pagesNeuronal Integration and CurcuitsOdyNo ratings yet
- Mrcs NotesDocument24 pagesMrcs Notesnob2011nob67% (3)