Download as pdf or txt
You might also like
- Biol 308 - Study QuestionsDocument47 pagesBiol 308 - Study QuestionsDivya100% (1)
- CPP by Sourav Kumar Giri PDFDocument133 pagesCPP by Sourav Kumar Giri PDFrednri0% (1)
- 2011 PipocDocument42 pages2011 PipocBung Harun0% (1)
- 2088 FullDocument17 pages2088 FullM.Febrian BachtiarNo ratings yet
- Gene Expressing FinalDocument14 pagesGene Expressing Finalwaliul islamNo ratings yet
- A Symphony of Transcription Factors For Gene ControlDocument19 pagesA Symphony of Transcription Factors For Gene ControlEdgardo Becerra BecerraNo ratings yet
- Bacterial Transcriptomics What Is Beyond The RNA Horiz-OmeDocument12 pagesBacterial Transcriptomics What Is Beyond The RNA Horiz-OmeIvaylo Nankov NaydenovNo ratings yet
- (17417899 - Reproduction) Regulation of Gene Transcription in The EpididymisDocument8 pages(17417899 - Reproduction) Regulation of Gene Transcription in The EpididymisAhmad SolihinNo ratings yet
- Functional Consequences of Developmentally Regulated Alternative SplicingDocument15 pagesFunctional Consequences of Developmentally Regulated Alternative SplicingAuinash KalsotraNo ratings yet
- Fis Upstream BindingDocument15 pagesFis Upstream BindingDiegoNo ratings yet
- A Unified Theory of Gene ExpressionDocument13 pagesA Unified Theory of Gene ExpressionManuel HernándezNo ratings yet
- D Mallika ComputerscienceDocument62 pagesD Mallika ComputerscienceDhilsanth SLNo ratings yet
- Wright 2015Document12 pagesWright 2015LasaroNo ratings yet
- The Plant Journal - 2004 - Zimmermann - Comprehensive Identification of Arabidopsis Thaliana MYB Transcription FactorsDocument13 pagesThe Plant Journal - 2004 - Zimmermann - Comprehensive Identification of Arabidopsis Thaliana MYB Transcription FactorsUhrigNo ratings yet
- Steroid Hor MonDocument14 pagesSteroid Hor MonchazmarilynNo ratings yet
- Transcription Factor Dynamics Mini ReviewDocument8 pagesTranscription Factor Dynamics Mini ReviewwebhappyNo ratings yet
- E2017005 PDFDocument6 pagesE2017005 PDFComputational and Structural Biotechnology JournalNo ratings yet
- Genome Res.-2012-Stadler-2418-26Document10 pagesGenome Res.-2012-Stadler-2418-26frostyNo ratings yet
- Transcription Sample Old Essay PDFDocument7 pagesTranscription Sample Old Essay PDFOccamsRazorNo ratings yet
- tmpE31A TMPDocument14 pagestmpE31A TMPFrontiersNo ratings yet
- Zhu GenomeRes2009 89yeastTFsDocument11 pagesZhu GenomeRes2009 89yeastTFsyp12345No ratings yet
- Epigenética y ARNDocument9 pagesEpigenética y ARNFrank BustosNo ratings yet
- The of Reporter Genes The Study of Microbial Ecology: Use inDocument12 pagesThe of Reporter Genes The Study of Microbial Ecology: Use inSadman RashidNo ratings yet
- Microreview: Quorum Sensing by Peptide Pheromones and Two-Component Signal-Transduction Systems in Gram-Positive BacteriaDocument10 pagesMicroreview: Quorum Sensing by Peptide Pheromones and Two-Component Signal-Transduction Systems in Gram-Positive BacteriaCristina AlexandraNo ratings yet
- Epigenetic Inheritance in PlantsDocument7 pagesEpigenetic Inheritance in Plantsapi-20009652No ratings yet
- Research ProposalDocument7 pagesResearch ProposalBitan BiswasNo ratings yet
- Journal Pcbi 1010087Document19 pagesJournal Pcbi 1010087rossi akalNo ratings yet
- Functionally Related Transcripts Have Common RNA MotifsDocument19 pagesFunctionally Related Transcripts Have Common RNA MotifsJuan Yujra CárdenasNo ratings yet
- Neurobiology of Pain: June Bryan de La Peña, Zachary T. Campbell TDocument6 pagesNeurobiology of Pain: June Bryan de La Peña, Zachary T. Campbell TPavithra MohanNo ratings yet
- Nihms 1025434Document12 pagesNihms 1025434Laura GarciaNo ratings yet
- Tmp38a4 TMPDocument9 pagesTmp38a4 TMPFrontiersNo ratings yet
- Bioinformatics: Discovery NoteDocument4 pagesBioinformatics: Discovery NotelaksiyerNo ratings yet
- sRNA 1Document8 pagessRNA 1Alex1546No ratings yet
- Review: A Decade of RiboswitchesDocument8 pagesReview: A Decade of RiboswitcheshansmeetNo ratings yet
- Molecular BiologyDocument9 pagesMolecular Biologyrifat RasheedNo ratings yet
- Ribo SwitchDocument10 pagesRibo Switchrag.1607No ratings yet
- Histone Deacetylases Transcriptional Control, and Cancer.Document16 pagesHistone Deacetylases Transcriptional Control, and Cancer.jose_gladiusNo ratings yet
- PQ 0403001990Document6 pagesPQ 0403001990Yuwono WibowoNo ratings yet
- Functional Enrichment Analysis Based On Long Noncoding RNA AssociationsDocument10 pagesFunctional Enrichment Analysis Based On Long Noncoding RNA AssociationsYamile A Rodríguez RiascosNo ratings yet
- Order ID 406406482 Gene ExpressionDocument4 pagesOrder ID 406406482 Gene ExpressionjohnessamburuNo ratings yet
- New EukDocument24 pagesNew EuknikhilsathwikNo ratings yet
- Capstone ReviewDocument28 pagesCapstone ReviewChad Kenneth CainNo ratings yet
- Castañeda 2016Document16 pagesCastañeda 2016papahojaloveNo ratings yet
- Gene Work FinalDocument5 pagesGene Work FinalpaurooshNo ratings yet
- Chloroplast Transformation For Engineering of PhotosynthesisDocument12 pagesChloroplast Transformation For Engineering of PhotosynthesisangkiNo ratings yet
- Nature 11247Document18 pagesNature 11247Cristina PavelNo ratings yet
- BT2030 NolDocument51 pagesBT2030 Nolkrishradhan1100% (1)
- 1 s2.0 S1535947621001584 MainDocument28 pages1 s2.0 S1535947621001584 MainBen DresimNo ratings yet
- EpigeneticsDocument5 pagesEpigeneticshurainsahar21No ratings yet
- Virus Res 2012Document12 pagesVirus Res 2012lubicanNo ratings yet
- Intrinsically Unstructured Proteins: Re-Assessing The Protein Structure-Function ParadigmDocument11 pagesIntrinsically Unstructured Proteins: Re-Assessing The Protein Structure-Function ParadigmJose DanielNo ratings yet
- List of Related AbstractsDocument5 pagesList of Related AbstractsAiman RoslizarNo ratings yet
- Evidence For The Association of Chromatin and Microrna Regulation in The Human GenomeDocument9 pagesEvidence For The Association of Chromatin and Microrna Regulation in The Human Genomeh234074No ratings yet
- Homeostasis in The Central Dogma of Molecular Biology: The Importance of mRNA InstabilityDocument23 pagesHomeostasis in The Central Dogma of Molecular Biology: The Importance of mRNA InstabilityAngelina KobanNo ratings yet
- Advances in Regulatory Network 2011Document15 pagesAdvances in Regulatory Network 2011nungkionNo ratings yet
- DNA Structure and Analysis: Chapter 14 (Translation and Protein)Document2 pagesDNA Structure and Analysis: Chapter 14 (Translation and Protein)Cobe Christian LascunaNo ratings yet
- Fisiologia Artigo1Document1 pageFisiologia Artigo1Jamylle CatharineNo ratings yet
- 13 Genes Dev. 2009 WiluszDocument12 pages13 Genes Dev. 2009 Wiluszmilenerato2240No ratings yet
- Dicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesDocument13 pagesDicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesAladár Pettkó-SzandtnerNo ratings yet
- Transduction Protein Interactions Define Specificity in Signal ProteinDocument22 pagesTransduction Protein Interactions Define Specificity in Signal ProteinnitishpathaniaNo ratings yet
- ChatGPT talks on science for young people: Molecular Biology!: Discover the secrets of life with the help of artificial intelligenceFrom EverandChatGPT talks on science for young people: Molecular Biology!: Discover the secrets of life with the help of artificial intelligenceNo ratings yet
- Transmembrane TransportersFrom EverandTransmembrane TransportersMichael W. QuickNo ratings yet
- Ajax & Javascript Libraries and Frameworks: Cs193C Summer 2015, YoungDocument4 pagesAjax & Javascript Libraries and Frameworks: Cs193C Summer 2015, YoungrednriNo ratings yet
- Algorithmic and Logical Thinking Development: Base of Programming SkillsDocument11 pagesAlgorithmic and Logical Thinking Development: Base of Programming SkillsrednriNo ratings yet
- The STRCHR Function: Using STRCHR To Search A String For A Single CharacterDocument20 pagesThe STRCHR Function: Using STRCHR To Search A String For A Single CharacterrednriNo ratings yet
- Hidden Markov Models in Bioinformatics: Example Domain: Gene FindingDocument32 pagesHidden Markov Models in Bioinformatics: Example Domain: Gene FindingrednriNo ratings yet
- A Brief Introduction To SAS Operators and FunctionsDocument7 pagesA Brief Introduction To SAS Operators and FunctionsrednriNo ratings yet
- Designing Voting Machines For Verification: N S T K D WDocument16 pagesDesigning Voting Machines For Verification: N S T K D WrednriNo ratings yet
- Finger Print Based Electronic Voting MachineDocument14 pagesFinger Print Based Electronic Voting MachinerednriNo ratings yet
- Ass 2 BioinformaticsDocument8 pagesAss 2 BioinformaticsMuhammad Anas JamshedNo ratings yet
- Genetics M ItDocument288 pagesGenetics M It007krunoNo ratings yet
- Biochemistry ReviewDocument13 pagesBiochemistry ReviewMohammad IzadiNo ratings yet
- Emetogenic Potential of Antineoplastic Agents PDFDocument1 pageEmetogenic Potential of Antineoplastic Agents PDFalfox2000No ratings yet
- Tools of BiotechnologyDocument10 pagesTools of Biotechnologyiamforu1No ratings yet
- What Happened To China Pharma Outsourcing Industry in 2008 - News CompileDocument55 pagesWhat Happened To China Pharma Outsourcing Industry in 2008 - News Compilejz9826100% (6)
- HPHDocument18 pagesHPHgopalc1940No ratings yet
- Biologie MolecularaDocument654 pagesBiologie MolecularaIulia Mihaela100% (2)
- Biotechnology Principles & Processes PDFDocument3 pagesBiotechnology Principles & Processes PDFMEGHA KRISHNANNo ratings yet
- Inventions/Discoveries Name of The Scientist/PersonDocument17 pagesInventions/Discoveries Name of The Scientist/PersonEngr Suleman MemonNo ratings yet
- Manufacturer Abbreviations : (An Abbott Company)Document11 pagesManufacturer Abbreviations : (An Abbott Company)Taylor McBrideNo ratings yet
- Production of Acetone-Butanol-ethanol From Spoilage Date PalmDocument7 pagesProduction of Acetone-Butanol-ethanol From Spoilage Date PalmI-Twins RgnNo ratings yet
- Top 100 Indian Pharma CompanyDocument3 pagesTop 100 Indian Pharma CompanyThavasi NadarNo ratings yet
- Biotech DirectoryDocument56 pagesBiotech DirectorySreesanth SaruvilNo ratings yet
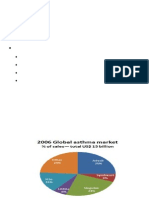
- Asthma MarketDocument18 pagesAsthma MarketPoojaNo ratings yet
- Applications of ZymographyDocument25 pagesApplications of ZymographyXênia SoutoNo ratings yet
- Ancaman Global Kesehatan Dan Finansial Di Era MDRODocument29 pagesAncaman Global Kesehatan Dan Finansial Di Era MDROYusuf AjaNo ratings yet
- 6.1 Cell Division 6.2 Cell Cycle and MitosisDocument57 pages6.1 Cell Division 6.2 Cell Cycle and Mitosiswickedbiology101No ratings yet
- Bet On One Big Idea-Or DiversifyDocument12 pagesBet On One Big Idea-Or Diversifyiris virtudezNo ratings yet
- Lec 6-10Document64 pagesLec 6-10shweta_agrawal24100% (1)
- Syllabus For M.S. (Pharm.) / PH.D Courses, Offered by NIPER - HyderabadDocument51 pagesSyllabus For M.S. (Pharm.) / PH.D Courses, Offered by NIPER - HyderabadAshok YadavNo ratings yet
- Metallomics and The Cell (Banci, Lucia)Document641 pagesMetallomics and The Cell (Banci, Lucia)ECienfuegos100% (1)
- BBL331 Course ContentDocument1 pageBBL331 Course ContentRoshineeNo ratings yet
- t4 SC 409 Genes and Inheritance Match and DrawDocument2 pagest4 SC 409 Genes and Inheritance Match and DrawNubar MammadovaNo ratings yet
- Pharmaceuticals Industry in IndiaDocument4 pagesPharmaceuticals Industry in IndiachetnaNo ratings yet
- Engem Eta Genetic ADocument8 pagesEngem Eta Genetic AgracyddNo ratings yet
- Bioinformatics Developments in IndiaDocument27 pagesBioinformatics Developments in IndiaBhupendra KhandelwalNo ratings yet
- Bosnian DNADocument8 pagesBosnian DNAJoe kNo ratings yet