Download as pdf or txt
You might also like
- Parts of A Bar.Document3 pagesParts of A Bar.MARY JOY VILLARUEL100% (1)
- National Geographic Instructor GuideDocument28 pagesNational Geographic Instructor GuideLuisOliveira100% (4)
- Oregano CapstoneDocument16 pagesOregano CapstonePrincess Cloewie Ferrer100% (1)
- Handbook Combined PDFDocument60 pagesHandbook Combined PDFHana Hill100% (2)
- Lu Et Al 2003 Sucesional Stages PDFDocument12 pagesLu Et Al 2003 Sucesional Stages PDFVanessa AraujoNo ratings yet
- Tree Species Composition, Richness and Diversity of The Mount Matutum Protected Landscape (MMPL), PhilippinesDocument12 pagesTree Species Composition, Richness and Diversity of The Mount Matutum Protected Landscape (MMPL), PhilippinesMd Ashikur RahmanNo ratings yet
- Costa Et Al. 2008 Gradients Within Gradients Central Amazonian ForestDocument11 pagesCosta Et Al. 2008 Gradients Within Gradients Central Amazonian ForestccruzvasconcelosNo ratings yet
- Vegetation Map - BINP - Babaasa Et Al. 2024Document13 pagesVegetation Map - BINP - Babaasa Et Al. 2024BabaasaNo ratings yet
- O62F3754Document216 pagesO62F3754jegarciapNo ratings yet
- Granato Souza2018Document12 pagesGranato Souza2018FressiaNo ratings yet
- Laurance Et Al - 2002 - ConBio PDFDocument14 pagesLaurance Et Al - 2002 - ConBio PDFMax Torres DíazNo ratings yet
- The Relevance of Geospatial Techniques in The Assessment of Forest Fragmentation of Anjaneri Hill, Nasik District, Maharashtra, IndiaDocument10 pagesThe Relevance of Geospatial Techniques in The Assessment of Forest Fragmentation of Anjaneri Hill, Nasik District, Maharashtra, IndiaIOSRjournalNo ratings yet
- Adjalla Et Al 2021, Soil Seed Bank Characteristics Along A Gradient of Past Human DisturbancesDocument13 pagesAdjalla Et Al 2021, Soil Seed Bank Characteristics Along A Gradient of Past Human DisturbancesADJALLANo ratings yet
- Fine-Scale - Habitat - Differentiation - Shapes - The - Comp ALICEDocument13 pagesFine-Scale - Habitat - Differentiation - Shapes - The - Comp ALICEluz marina mulato campoNo ratings yet
- Forest Fragmentation and Its Impact On Species Diversity: An Analysis Using Remote Sensing and GISDocument18 pagesForest Fragmentation and Its Impact On Species Diversity: An Analysis Using Remote Sensing and GISrshNo ratings yet
- Species Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringDocument13 pagesSpecies Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringLight WhiteNo ratings yet
- House 2003Document16 pagesHouse 2003sandrino1091No ratings yet
- Foresr Health MonitoringDocument15 pagesForesr Health MonitoringMaulana Ikhsan HasibuanNo ratings yet
- Deforestation in Heilongjiang Province of China, 1896e2000 Severity, PDFDocument8 pagesDeforestation in Heilongjiang Province of China, 1896e2000 Severity, PDFAnamaria BăltuțăNo ratings yet
- Small Mammal Assemblages in A Disturbed Tropical LDocument10 pagesSmall Mammal Assemblages in A Disturbed Tropical LHeiner SaldañaNo ratings yet
- Roças Et Al 2001Document8 pagesRoças Et Al 2001Sabrina QueirozNo ratings yet
- 10 - 2017 - Melo Júnior Et Al. 2017 - Ciência & NaturaDocument10 pages10 - 2017 - Melo Júnior Et Al. 2017 - Ciência & NaturaÍgor Abba ArriolaNo ratings yet
- Frontiers in Ecology and The Environment - 2023Document9 pagesFrontiers in Ecology and The Environment - 2023Igor OliveiraNo ratings yet
- Jadan Et Al 2022 Forest EcosystemsDocument9 pagesJadan Et Al 2022 Forest EcosystemsOswaldo JadanNo ratings yet
- NascimentoDocument11 pagesNascimentotaucci123No ratings yet
- Stancioiu-OHara2006 Article RegenerationGrowthInDifferentLDocument12 pagesStancioiu-OHara2006 Article RegenerationGrowthInDifferentLBroXNo ratings yet
- Aparecido2019 - SapwoodDocument15 pagesAparecido2019 - SapwoodcristineNo ratings yet
- Association For Tropical Biology and Conservation BiotropicaDocument11 pagesAssociation For Tropical Biology and Conservation BiotropicazaydaNo ratings yet
- Anthropogenic Disturbances and Plant Diversity of The Madhupur Sal Forests (Shorea Robusta C.F. Gaertn) of BangladeshDocument13 pagesAnthropogenic Disturbances and Plant Diversity of The Madhupur Sal Forests (Shorea Robusta C.F. Gaertn) of BangladeshMonabbirNo ratings yet
- Uezu, Beyer and Metzger 2008Document16 pagesUezu, Beyer and Metzger 2008Vitoria Duarte DerissoNo ratings yet
- SEv2 c23Document14 pagesSEv2 c23Anonymous BkmsKXzwyKNo ratings yet
- tmp69 TMPDocument12 pagestmp69 TMPFrontiersNo ratings yet
- (Q1) Gholami2017Document8 pages(Q1) Gholami2017Salma FahimaNo ratings yet
- WernerEtAl 2011 IsTheResilienceOfEpiphyteAssemblagesDocument6 pagesWernerEtAl 2011 IsTheResilienceOfEpiphyteAssemblagesLau Rodríguez LavínNo ratings yet
- 1-Zuquimetal2012 ConverisDocument40 pages1-Zuquimetal2012 ConverisHeriberto Pinzón GómezNo ratings yet
- A Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et AlDocument20 pagesA Comparison of The Wood Anatomy of 11 Species From Two Cerrado Habitats (Cerrado S.S. and Adjacent Gallery Forest) - Sonsin Et Allihelih1No ratings yet
- Santos Et Al, 2008 - Drastic Erosion in Functional Attributes of Tree Assemblagesin Atlantic Forest Fragments of Northeastern BrazilDocument12 pagesSantos Et Al, 2008 - Drastic Erosion in Functional Attributes of Tree Assemblagesin Atlantic Forest Fragments of Northeastern BrazilMarcondes OliveiraNo ratings yet
- Forest Effect On Seed Bank 2017Document8 pagesForest Effect On Seed Bank 2017Khalil AliNo ratings yet
- 464 FullDocument11 pages464 FullLucía SolerNo ratings yet
- Assessing Impacts of Selective Logging On Water, Energy, and CarbonDocument30 pagesAssessing Impacts of Selective Logging On Water, Energy, and CarbonDênis Aguero do NascimentoNo ratings yet
- Neotropical Rainforest Restoration, Comparing Passive, Plantation and Nucleation ApproachesDocument14 pagesNeotropical Rainforest Restoration, Comparing Passive, Plantation and Nucleation ApproachesMirabelNo ratings yet
- Tuomisto Et Al 2003 - ScienceDocument5 pagesTuomisto Et Al 2003 - ScienceKátia FernandaNo ratings yet
- Vegetation Dynamics and Soil Nutrients Across Different Shifting CultivationDocument8 pagesVegetation Dynamics and Soil Nutrients Across Different Shifting CultivationReni Setyo WahyuningtyasNo ratings yet
- Aldrich 1998Document12 pagesAldrich 1998Rany sanchium tawamNo ratings yet
- Determining Aboveground Biomass of The Forest Successional Chronosequence in A Test-Site of Brazilian Amazon Through X-And L - Band Data AnalysisDocument10 pagesDetermining Aboveground Biomass of The Forest Successional Chronosequence in A Test-Site of Brazilian Amazon Through X-And L - Band Data AnalysisJeanpierre RodríguezNo ratings yet
- Urbina-Cardona Et Al 2006 BiolconsDocument15 pagesUrbina-Cardona Et Al 2006 BiolconsDaniel VásquezNo ratings yet
- Review Article Influence of Climatic Variables, Forest Type, and Condition On Activity Patterns of Geoffroyi's Spider Monkeys Throughout MesoamericaDocument10 pagesReview Article Influence of Climatic Variables, Forest Type, and Condition On Activity Patterns of Geoffroyi's Spider Monkeys Throughout MesoamericaFreakyRustlee LeoragNo ratings yet
- Duque Et Al. 2017 Insights Into Regional Patterns of Amazonian ForestDocument18 pagesDuque Et Al. 2017 Insights Into Regional Patterns of Amazonian ForestccruzvasconcelosNo ratings yet
- Pike Etal 2001 PanamaDocument14 pagesPike Etal 2001 PanamaAnonymous ek4z9UBNo ratings yet
- Stand Structure of Pasoh Forest Reserve, A Lowland Rain Forest in Peninsular MalaysiaDocument11 pagesStand Structure of Pasoh Forest Reserve, A Lowland Rain Forest in Peninsular MalaysiaKung FooNo ratings yet
- Dispersal Traits Determine Passive Restoration Trajectory of A Nigerian Montane ForestDocument9 pagesDispersal Traits Determine Passive Restoration Trajectory of A Nigerian Montane ForestsebastianNo ratings yet
- Growth-Ring Boundary Anatomy and Dendrochronological Potential in A Moist Tropical Forest in Northeastern BangladeshDocument19 pagesGrowth-Ring Boundary Anatomy and Dendrochronological Potential in A Moist Tropical Forest in Northeastern BangladeshAnthony GuerraNo ratings yet
- Muñoz Mazón Et Al., 2020Document9 pagesMuñoz Mazón Et Al., 2020azeta zetaNo ratings yet
- Importance of Structure and Its Measurement in Quantifying Function of Forest EcosystemsDocument16 pagesImportance of Structure and Its Measurement in Quantifying Function of Forest EcosystemsRodrigo vazquez moralesNo ratings yet
- Photosynthesis of Nine Pioneer Borneo in Relation To Life HistoryDocument17 pagesPhotosynthesis of Nine Pioneer Borneo in Relation To Life HistoryRachman FamieNo ratings yet
- American J Primatol - 2020 - Steffens - Occurrence of Lemurs in Landscapes and Their Species Specific Scale Responses ToDocument9 pagesAmerican J Primatol - 2020 - Steffens - Occurrence of Lemurs in Landscapes and Their Species Specific Scale Responses ToDellNo ratings yet
- Asquith y Mejia - Chang-Mammals, Edge Effects, and The Loss of Tropical Forest Diversity-Ecology-2005Document13 pagesAsquith y Mejia - Chang-Mammals, Edge Effects, and The Loss of Tropical Forest Diversity-Ecology-2005braulioasensioNo ratings yet
- Landscape Effects On Taxonomic and Functional Diversity of Dung Beetle Assemblages in A Highly Fragmented Tropical ForestDocument10 pagesLandscape Effects On Taxonomic and Functional Diversity of Dung Beetle Assemblages in A Highly Fragmented Tropical ForestCarlos Elias Altamar BolibarNo ratings yet
- Composition, Structure and Diversity of Tree Species Along An Elevational Gradient in Jammu Province of North-Western Himalayas, Jammu and Kashmir, IndiaDocument12 pagesComposition, Structure and Diversity of Tree Species Along An Elevational Gradient in Jammu Province of North-Western Himalayas, Jammu and Kashmir, IndiaInternational Network For Natural Sciences100% (1)
- Puker 24Document13 pagesPuker 24Debora Lima SantosNo ratings yet
- A Word of Caution When Planning Forest Management Using Projections of Tree Species Range ShiftsDocument13 pagesA Word of Caution When Planning Forest Management Using Projections of Tree Species Range ShiftsjuanbvNo ratings yet
- Remotesensing 05 05493 v3Document21 pagesRemotesensing 05 05493 v3fabricio alvesNo ratings yet
- Detailed Maps of Tropical Forest Types Are Within Reach: Forest Tree Communities For Trinidad and TobagoDocument20 pagesDetailed Maps of Tropical Forest Types Are Within Reach: Forest Tree Communities For Trinidad and TobagoCarlos GrupotecNo ratings yet
- Ecology and Sustainability of Southern Temperate EcosystemsFrom EverandEcology and Sustainability of Southern Temperate EcosystemsTW NortonNo ratings yet
- Press Release Coty Closing Hypermarcas AcquisitionDocument2 pagesPress Release Coty Closing Hypermarcas AcquisitionTytyt TyyNo ratings yet
- Transboundary HazeDocument15 pagesTransboundary HazeTytyt TyyNo ratings yet
- Singapore Affected by Severe Transboundary Smoke Haze in June 2013Document5 pagesSingapore Affected by Severe Transboundary Smoke Haze in June 2013Tytyt TyyNo ratings yet
- Haze and Health EffectsDocument5 pagesHaze and Health EffectsTytyt TyyNo ratings yet
- 2014 Americans Overseas Ballot Mailing DeadlinesDocument7 pages2014 Americans Overseas Ballot Mailing DeadlinesTytyt TyyNo ratings yet
- Haze 2013Document5 pagesHaze 2013Tytyt TyyNo ratings yet
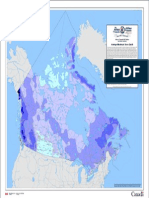
- Average Maximum Snow Depth: Atlas of Canada 6th Edition (Archival Version)Document1 pageAverage Maximum Snow Depth: Atlas of Canada 6th Edition (Archival Version)Tytyt TyyNo ratings yet
- The Real Story Behind The: South China Sea DisputeDocument2 pagesThe Real Story Behind The: South China Sea DisputeTytyt TyyNo ratings yet
- Lalita BoonpriwanDocument17 pagesLalita BoonpriwanTytyt TyyNo ratings yet
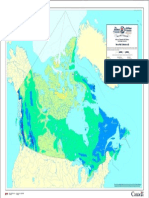
- Snowfall (Nunavut) : Atlas of Canada 6th Edition (Archival Version)Document1 pageSnowfall (Nunavut) : Atlas of Canada 6th Edition (Archival Version)Tytyt TyyNo ratings yet
- Ipcc Far WG II Chapter 07Document37 pagesIpcc Far WG II Chapter 07Tytyt TyyNo ratings yet
- LSM3254 TestDocument1 pageLSM3254 TestTytyt TyyNo ratings yet
- St. John The Devine Sermon Finakd) DumorDocument9 pagesSt. John The Devine Sermon Finakd) DumorTytyt TyyNo ratings yet
- Agriculture Management Approaches Pankaj Kumar Saraswat Full ChapterDocument51 pagesAgriculture Management Approaches Pankaj Kumar Saraswat Full Chapterhomer.griffin661100% (9)
- Eyelids Eyelids: 7 Semester Dr. Monika Mahat 3 Year Resident Ophthalmology Department LMCTHDocument64 pagesEyelids Eyelids: 7 Semester Dr. Monika Mahat 3 Year Resident Ophthalmology Department LMCTHSubash BasnetNo ratings yet
- Technical Data Sheet: Automotive Aftermarket Division 3M™ 09376 Perfect-It III Machine PolishDocument3 pagesTechnical Data Sheet: Automotive Aftermarket Division 3M™ 09376 Perfect-It III Machine Polishm_a_1993No ratings yet
- Delivery, Inspection and Quality Assurance: Learning Outcomes (Co1-Po4)Document14 pagesDelivery, Inspection and Quality Assurance: Learning Outcomes (Co1-Po4)nadewhoNo ratings yet
- Safety Standard For Horizontal Carousel Material Handling and Associated EquipmentDocument19 pagesSafety Standard For Horizontal Carousel Material Handling and Associated EquipmentIntegrated Systems DesignNo ratings yet
- Regla de Ergonomía 1Document15 pagesRegla de Ergonomía 1Valeria RincónNo ratings yet
- Simplest Ganapathi HomaDocument4 pagesSimplest Ganapathi HomaAnand AbheyNo ratings yet
- Asymmetric Dual N-Channel Trench MOSFET 30V: General Description FeaturesDocument9 pagesAsymmetric Dual N-Channel Trench MOSFET 30V: General Description Featureschristian romoNo ratings yet
- s555qw5w60101011010 Scalloppsnskskdndndn D Scalllpssond Sjsjsns S 3u373636191ir827472y152ta D D. DJDNDJFNFNF Ala. K K J N K nk755555511112727273737 11515152t2cawuwuwu272727 1116263636bllop - 1Document4 pagess555qw5w60101011010 Scalloppsnskskdndndn D Scalllpssond Sjsjsns S 3u373636191ir827472y152ta D D. DJDNDJFNFNF Ala. K K J N K nk755555511112727273737 11515152t2cawuwuwu272727 1116263636bllop - 1Red DiggerNo ratings yet
- Social InfluenceDocument26 pagesSocial Influenceattique100% (2)
- Sludge TreatmentDocument34 pagesSludge TreatmentNurul Fatihah SyazwaniNo ratings yet
- Heat Exchanger Types Spring 2018 PDFDocument26 pagesHeat Exchanger Types Spring 2018 PDFAmeer AlfuqahaNo ratings yet
- Specification Chart of Pipes - Stainless Steel - Seamless Pipe - Mild Steel - Hollow SteelDocument5 pagesSpecification Chart of Pipes - Stainless Steel - Seamless Pipe - Mild Steel - Hollow SteelSarita BoteNo ratings yet
- Boards - OmaeDocument4 pagesBoards - OmaeJoyce EbolNo ratings yet
- Model Element Table - LOD MATRIX INSTRUCTIONSDocument6 pagesModel Element Table - LOD MATRIX INSTRUCTIONSMark ChuoNo ratings yet
- Journal Pre-Proof: Clinical Epidemiology and Global HealthDocument17 pagesJournal Pre-Proof: Clinical Epidemiology and Global HealthYurisma KhoerulNo ratings yet
- T27 T11 (SIeve ANalysis)Document46 pagesT27 T11 (SIeve ANalysis)Don Procopio IINo ratings yet
- Environmental Problems WorksheetDocument2 pagesEnvironmental Problems WorksheetAnnamária Polacsek100% (1)
- HRDM 11-7 - WEEK 06 To 07 - Activity and Output - Ma. Christine C. MontejoDocument13 pagesHRDM 11-7 - WEEK 06 To 07 - Activity and Output - Ma. Christine C. Montejochristine montejoNo ratings yet
- Project Standard Specification: Packaged Cooling Towers 15640 - Page 1/9Document9 pagesProject Standard Specification: Packaged Cooling Towers 15640 - Page 1/9adel rihanaNo ratings yet
- The Organization of Modern KitchensDocument9 pagesThe Organization of Modern KitchensJohnson Macayan FernándezNo ratings yet
- Chapter 3Document39 pagesChapter 3amiranurul50% (2)
- Here There Be Tygers by Stephen KingDocument3 pagesHere There Be Tygers by Stephen KingxXPizzaPresidentXx100% (1)
- Pharmacology Powerpoint Group 1Document38 pagesPharmacology Powerpoint Group 1Daniel LaurenteNo ratings yet
- Red Cross Red Crescent Magazine, No. 2, 2013Document32 pagesRed Cross Red Crescent Magazine, No. 2, 2013International Committee of the Red Cross100% (1)
- Personal Finance Canadian 4th Edition Kapoor Test BankDocument30 pagesPersonal Finance Canadian 4th Edition Kapoor Test Bankkhuyenfrederickgjk8100% (35)