
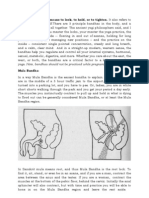
BJ 2002 Graton Mantulin Glaser
BJ 2002 Graton Mantulin Glaser
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cure Any Disease BookDocument29 pagesCure Any Disease Bookadina_bluebee100% (5)
- Mula Uddiyan Jalandhar Bandha & Maha MudraDocument4 pagesMula Uddiyan Jalandhar Bandha & Maha MudraBhavesh1801No ratings yet
- Education Support Pack Special SchoolsDocument144 pagesEducation Support Pack Special SchoolsRameeSahibaNo ratings yet
- Jurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootDocument6 pagesJurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootAchmad Zam Zam AghazyNo ratings yet
- Anders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFDocument433 pagesAnders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFPhil BoileauNo ratings yet
- Polygenic InheritanceDocument45 pagesPolygenic Inheritanceapi-19916399No ratings yet
- Webb Juro Theondre R. Mirondo Gen Bio Written Work #2Document5 pagesWebb Juro Theondre R. Mirondo Gen Bio Written Work #2Mirondo Webb Juro TheondreNo ratings yet
- Bio Chap 8 Circulatory SystemDocument11 pagesBio Chap 8 Circulatory SystemJyoti AmbwaniNo ratings yet
- Cell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNADocument8 pagesCell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNAQadirNo ratings yet
- Unit 5 - Topic 4 PathologyDocument9 pagesUnit 5 - Topic 4 PathologyHùng Mạnh NguyễnNo ratings yet
- Chapter 29 Child With CancerDocument9 pagesChapter 29 Child With CancerMARCERA JERALDINE ALESSA P.No ratings yet
- Bloodbld 2020009530Document16 pagesBloodbld 2020009530Yanet FrancoNo ratings yet
- Raw Food DilemmasDocument11 pagesRaw Food Dilemmas90002No ratings yet
- BunyaviridaeDocument12 pagesBunyaviridaeRia AlcantaraNo ratings yet
- Report STS MH 4 6 - 1stsem23 24Document3 pagesReport STS MH 4 6 - 1stsem23 24Kurt SecretarioNo ratings yet
- Uric Acid, Blood Pressure, and Gout Management (PDFDrive)Document200 pagesUric Acid, Blood Pressure, and Gout Management (PDFDrive)Aurveda SE HealNo ratings yet
- EUPO2013 Coursebook OnlineDocument220 pagesEUPO2013 Coursebook Onlineiuliad88100% (1)
- Disadvantages of Inorganic FoodDocument12 pagesDisadvantages of Inorganic FoodYatharth Sahu100% (1)
- What Causes IBS?Document4 pagesWhat Causes IBS?prima suci angrainiNo ratings yet
- L9-10: Genetic Engineering of Biosynthetic Pathways in Transgenic PlantsDocument72 pagesL9-10: Genetic Engineering of Biosynthetic Pathways in Transgenic PlantsTing Yan LeeNo ratings yet
- Epilepsia - 2022 - Hirsch - ILAE Definition of The Idiopathic Generalized Epilepsy Syndromes Position Statement by TheDocument25 pagesEpilepsia - 2022 - Hirsch - ILAE Definition of The Idiopathic Generalized Epilepsy Syndromes Position Statement by TheIngrid RivadeneiraNo ratings yet
- Atopic Dermatitis and HomoeopathyDocument8 pagesAtopic Dermatitis and HomoeopathyDr. Rajneesh Kumar Sharma MD Hom100% (2)
- Topic Test - Genetics MemoDocument5 pagesTopic Test - Genetics Memomaiselatebogo4No ratings yet
- Botany (S)Document11 pagesBotany (S)Aniket KhargaNo ratings yet
- COVID-19 Information: Ncbi S Kip To Main Content S Kip To Navigation Resources How To A Bout NCBI AccesskeysDocument5 pagesCOVID-19 Information: Ncbi S Kip To Main Content S Kip To Navigation Resources How To A Bout NCBI AccesskeyscatheirneNo ratings yet
- Nursing Management in Cancer CareDocument27 pagesNursing Management in Cancer CareFev BanataoNo ratings yet
- Phylogenetic and Phytochemical Characteristics Ch8 KLEINDocument22 pagesPhylogenetic and Phytochemical Characteristics Ch8 KLEINrobynkleinNo ratings yet
- Renal Cell Carcinoma: Diagnosis, Staging, and SurveillanceDocument18 pagesRenal Cell Carcinoma: Diagnosis, Staging, and SurveillanceMutia IsanaNo ratings yet
- Jof 07 00844 v2Document11 pagesJof 07 00844 v2Sonia HernándezNo ratings yet

















































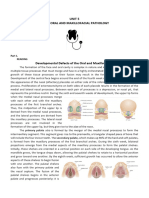



















Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5820)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cure Any Disease BookDocument29 pagesCure Any Disease Bookadina_bluebee100% (5)
- Mula Uddiyan Jalandhar Bandha & Maha MudraDocument4 pagesMula Uddiyan Jalandhar Bandha & Maha MudraBhavesh1801No ratings yet
- Education Support Pack Special SchoolsDocument144 pagesEducation Support Pack Special SchoolsRameeSahibaNo ratings yet
- Jurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootDocument6 pagesJurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootAchmad Zam Zam AghazyNo ratings yet
- Anders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFDocument433 pagesAnders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFPhil BoileauNo ratings yet
- Polygenic InheritanceDocument45 pagesPolygenic Inheritanceapi-19916399No ratings yet
- Webb Juro Theondre R. Mirondo Gen Bio Written Work #2Document5 pagesWebb Juro Theondre R. Mirondo Gen Bio Written Work #2Mirondo Webb Juro TheondreNo ratings yet
- Bio Chap 8 Circulatory SystemDocument11 pagesBio Chap 8 Circulatory SystemJyoti AmbwaniNo ratings yet
- Cell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNADocument8 pagesCell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNAQadirNo ratings yet
- Unit 5 - Topic 4 PathologyDocument9 pagesUnit 5 - Topic 4 PathologyHùng Mạnh NguyễnNo ratings yet
- Chapter 29 Child With CancerDocument9 pagesChapter 29 Child With CancerMARCERA JERALDINE ALESSA P.No ratings yet
- Bloodbld 2020009530Document16 pagesBloodbld 2020009530Yanet FrancoNo ratings yet
- Raw Food DilemmasDocument11 pagesRaw Food Dilemmas90002No ratings yet
- BunyaviridaeDocument12 pagesBunyaviridaeRia AlcantaraNo ratings yet
- Report STS MH 4 6 - 1stsem23 24Document3 pagesReport STS MH 4 6 - 1stsem23 24Kurt SecretarioNo ratings yet
- Uric Acid, Blood Pressure, and Gout Management (PDFDrive)Document200 pagesUric Acid, Blood Pressure, and Gout Management (PDFDrive)Aurveda SE HealNo ratings yet
- EUPO2013 Coursebook OnlineDocument220 pagesEUPO2013 Coursebook Onlineiuliad88100% (1)
- Disadvantages of Inorganic FoodDocument12 pagesDisadvantages of Inorganic FoodYatharth Sahu100% (1)
- What Causes IBS?Document4 pagesWhat Causes IBS?prima suci angrainiNo ratings yet
- L9-10: Genetic Engineering of Biosynthetic Pathways in Transgenic PlantsDocument72 pagesL9-10: Genetic Engineering of Biosynthetic Pathways in Transgenic PlantsTing Yan LeeNo ratings yet
- Epilepsia - 2022 - Hirsch - ILAE Definition of The Idiopathic Generalized Epilepsy Syndromes Position Statement by TheDocument25 pagesEpilepsia - 2022 - Hirsch - ILAE Definition of The Idiopathic Generalized Epilepsy Syndromes Position Statement by TheIngrid RivadeneiraNo ratings yet
- Atopic Dermatitis and HomoeopathyDocument8 pagesAtopic Dermatitis and HomoeopathyDr. Rajneesh Kumar Sharma MD Hom100% (2)
- Topic Test - Genetics MemoDocument5 pagesTopic Test - Genetics Memomaiselatebogo4No ratings yet
- Botany (S)Document11 pagesBotany (S)Aniket KhargaNo ratings yet
- COVID-19 Information: Ncbi S Kip To Main Content S Kip To Navigation Resources How To A Bout NCBI AccesskeysDocument5 pagesCOVID-19 Information: Ncbi S Kip To Main Content S Kip To Navigation Resources How To A Bout NCBI AccesskeyscatheirneNo ratings yet
- Nursing Management in Cancer CareDocument27 pagesNursing Management in Cancer CareFev BanataoNo ratings yet
- Phylogenetic and Phytochemical Characteristics Ch8 KLEINDocument22 pagesPhylogenetic and Phytochemical Characteristics Ch8 KLEINrobynkleinNo ratings yet
- Renal Cell Carcinoma: Diagnosis, Staging, and SurveillanceDocument18 pagesRenal Cell Carcinoma: Diagnosis, Staging, and SurveillanceMutia IsanaNo ratings yet
- Jof 07 00844 v2Document11 pagesJof 07 00844 v2Sonia HernándezNo ratings yet