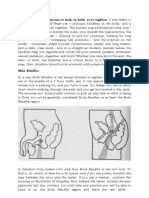
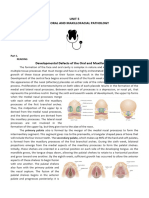
Download as pdf or txt
You might also like
- Oet ReadingDocument15 pagesOet Readingnj0% (1)
- Cure Any Disease BookDocument29 pagesCure Any Disease Bookadina_bluebee100% (5)
- Abe7729 Kienle SMDocument24 pagesAbe7729 Kienle SMIntan Yulia SafarinaNo ratings yet
- Cross 1987 Catecholamines LympohcytesDocument7 pagesCross 1987 Catecholamines LympohcytesMartin BadovNo ratings yet
- Chung SMDocument27 pagesChung SMNurul HidayahNo ratings yet
- Effet Glucane Reishi 19 Janv 2023Document8 pagesEffet Glucane Reishi 19 Janv 2023ilyNo ratings yet
- 9747-Supplementary Appendix-74841-1-10-20210329 PDFDocument19 pages9747-Supplementary Appendix-74841-1-10-20210329 PDFTianliang GuoNo ratings yet
- MMC 1Document8 pagesMMC 1anower jabedNo ratings yet
- 1 s2.0 S0014482702955813 MainDocument12 pages1 s2.0 S0014482702955813 MainKevin MaiseyNo ratings yet
- Antitumor Effect of RGD-4C-GG - (KLAKLAK) Peptide in Mouse B16 MELANOMADocument5 pagesAntitumor Effect of RGD-4C-GG - (KLAKLAK) Peptide in Mouse B16 MELANOMAacbgdvNo ratings yet
- Cisplatin Cell Cicle ArrestDocument7 pagesCisplatin Cell Cicle ArrestGabriele KrauseNo ratings yet
- Pnas 201311249siDocument9 pagesPnas 201311249sigenpercraftNo ratings yet
- Dodson MCBDocument12 pagesDodson MCBRonen ShemeshNo ratings yet
- Myc DownregulationDocument10 pagesMyc Downregulationkdcmbmwg9pNo ratings yet
- Nm.2365 SupplementsDocument12 pagesNm.2365 SupplementsdavdavdavdavdavdavdaNo ratings yet
- tmp37AD TMPDocument11 pagestmp37AD TMPFrontiersNo ratings yet
- Stage 1: Mesoendoderm: Supplemental Data Research Design and Methods Differentiation of Human ES CellsDocument10 pagesStage 1: Mesoendoderm: Supplemental Data Research Design and Methods Differentiation of Human ES CellsNyna KawlesNo ratings yet
- Quercetin Inhibits Hydrogen Peroxide (H O) - Induced Nf-Binding Activity and Dna Damage in Hepg2 CellsDocument7 pagesQuercetin Inhibits Hydrogen Peroxide (H O) - Induced Nf-Binding Activity and Dna Damage in Hepg2 CellsAnonymous wbRUEuDNo ratings yet
- CPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerDocument10 pagesCPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerFerdina NidyasariNo ratings yet
- The Tgf-B-Induced Up-Regulation of Nkg2Dls Requires Akt/Gsk-3 B-Mediated Stabilization of Sp1Document11 pagesThe Tgf-B-Induced Up-Regulation of Nkg2Dls Requires Akt/Gsk-3 B-Mediated Stabilization of Sp1João AndradeNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsSergeat18BNo ratings yet
- A Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLDocument8 pagesA Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLTareeqanwar MohammedNo ratings yet
- Ijbsv 18 P 3470 S 1Document15 pagesIjbsv 18 P 3470 S 1李秀赫No ratings yet
- mRNA Released From Astrocytes Trough Mixrovesicles Regulates Dendritic GrowthDocument21 pagesmRNA Released From Astrocytes Trough Mixrovesicles Regulates Dendritic GrowthDiego GaeteNo ratings yet
- Supplemental Information The Neuronal Repellent SLIT2 Is A Target For Repression by EZH2 in Prostate CancerDocument11 pagesSupplemental Information The Neuronal Repellent SLIT2 Is A Target For Repression by EZH2 in Prostate Cancermicrobotica126No ratings yet
- Int. Immunol.-1999-Staal-317-23Document7 pagesInt. Immunol.-1999-Staal-317-23Karthik AripakaNo ratings yet
- MCB00436-22-Merged - PDF MergedDocument47 pagesMCB00436-22-Merged - PDF MergedHector Ivan Saldivar CeronNo ratings yet
- Loreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingDocument15 pagesLoreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingAnadahiNo ratings yet
- pnas.201111575SIDocument4 pagespnas.201111575SIAlain Marc GuillemNo ratings yet
- Adiliaghdam Et Al. 2022 - Sciimmunol - Abn6660 - SMDocument26 pagesAdiliaghdam Et Al. 2022 - Sciimmunol - Abn6660 - SMAchille BroggiNo ratings yet
- 1996 - Determination of Life-Span in Caenorhabditis Elegans by Four Clock GenesDocument5 pages1996 - Determination of Life-Span in Caenorhabditis Elegans by Four Clock GenesRaymond LaBoyNo ratings yet
- Rapid: PublicationDocument6 pagesRapid: PublicationRaphael Fontoura OliveiraNo ratings yet
- Refolding and Characterization of RecombDocument6 pagesRefolding and Characterization of RecombMobarakNo ratings yet
- Shen 2018Document6 pagesShen 2018Arturo JiménezNo ratings yet
- Sun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDocument7 pagesSun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDaniela Patricia Alvarez AravenaNo ratings yet
- Duration of Mitotic CellDocument20 pagesDuration of Mitotic CellHà Thy Nguyễn ĐỗNo ratings yet
- Paper Target Validation Bump and HoleDocument7 pagesPaper Target Validation Bump and HolemarylenedemanNo ratings yet
- The Sympathetic Nervous System Modulates CD4+FoxP3+ Regulatory T Cells Via A TGFDocument9 pagesThe Sympathetic Nervous System Modulates CD4+FoxP3+ Regulatory T Cells Via A TGFPedro FonsecaNo ratings yet
- Pnas 201717789siDocument11 pagesPnas 201717789siygilad9139No ratings yet
- Tumor Necrosis Factor A Stimulates Human Clara Cell Secretory Protein Production by Human Airway Epithelial Cells.1998Document7 pagesTumor Necrosis Factor A Stimulates Human Clara Cell Secretory Protein Production by Human Airway Epithelial Cells.1998Fina Herlinda NurNo ratings yet
- Protective Effect of Antioxidant Enzymes Against Drug Cytotoxicity in Mcf-7 CellsDocument3 pagesProtective Effect of Antioxidant Enzymes Against Drug Cytotoxicity in Mcf-7 CellsVidhi AdilNo ratings yet
- Document 1Document7 pagesDocument 1xaero1234No ratings yet
- Nabel Et Al 2007 Lysis of Human Immunodeficiency Virus Type 1 by A Specific Secreted Human Phospholipase A2Document7 pagesNabel Et Al 2007 Lysis of Human Immunodeficiency Virus Type 1 by A Specific Secreted Human Phospholipase A2Marry FCCNo ratings yet
- Fan2016 Tissue Repair and Regeneration PDFDocument15 pagesFan2016 Tissue Repair and Regeneration PDFDavid LuNo ratings yet
- Said AhmedDocument19 pagesSaid Ahmedahmed mahdiNo ratings yet
- Adamiak Et Al. - 2018 - Induced Pluripotent Stem Cell (iPSC) - Derived Extracellular Vesicles Are Safer and More Effective For Cardiac RepDocument33 pagesAdamiak Et Al. - 2018 - Induced Pluripotent Stem Cell (iPSC) - Derived Extracellular Vesicles Are Safer and More Effective For Cardiac Repscribd mendeleyNo ratings yet
- Pnas 2304319120 SappDocument15 pagesPnas 2304319120 SappfilymascoloNo ratings yet
- Protein Phosphatase 2C (PP2C) Is Responsible For VP-induced Dephosphorylation of AQP2 Serine 261Document40 pagesProtein Phosphatase 2C (PP2C) Is Responsible For VP-induced Dephosphorylation of AQP2 Serine 261Almas TNo ratings yet
- Pic RenderDocument9 pagesPic Renderkora_c995No ratings yet
- NIHMS506822 Supplement 01Document18 pagesNIHMS506822 Supplement 01Bingyu XieNo ratings yet
- Immunology - 2008 - Montcuquet - Regulatory T Cell Expansion and Function Do Not Account For The Impaired Alloreactivity ofDocument11 pagesImmunology - 2008 - Montcuquet - Regulatory T Cell Expansion and Function Do Not Account For The Impaired Alloreactivity ofciara babyNo ratings yet
- DUAN (2019) LncRNA TUG1 Aggravates The Progression of Cervical Cancer by Binding PUM2Document8 pagesDUAN (2019) LncRNA TUG1 Aggravates The Progression of Cervical Cancer by Binding PUM2Arissa KohataNo ratings yet
- A Three-Symbol Code For Organized Proteomes Based On Cyclical Imaging of Protein LocationsDocument9 pagesA Three-Symbol Code For Organized Proteomes Based On Cyclical Imaging of Protein LocationsJulio YarNo ratings yet
- PUBLICADO JCBiochem - 2012Document12 pagesPUBLICADO JCBiochem - 2012Claudia BuitragoNo ratings yet
- Appearance of Water Channels in Xenopus Oocytes Expressing Red Cell CHIP28 ProteinDocument3 pagesAppearance of Water Channels in Xenopus Oocytes Expressing Red Cell CHIP28 ProteinKavi JainNo ratings yet
- Reversal of Streptozotocin-Induced Hyperglycemia by Transplantation of Pseudoislets Consisting of From Ductal CellsDocument6 pagesReversal of Streptozotocin-Induced Hyperglycemia by Transplantation of Pseudoislets Consisting of From Ductal CellskalwusaidiNo ratings yet
- Boumediene Bouzahzah - 2001 - 274 PDFDocument15 pagesBoumediene Bouzahzah - 2001 - 274 PDFarnipahlawaniNo ratings yet
- 16 - 2021 - Vieira-Neto Et Al. Effect of Source and Amount of Vitamin D On Function and MRNA Expression in Immune Cells in Dairy CowsDocument16 pages16 - 2021 - Vieira-Neto Et Al. Effect of Source and Amount of Vitamin D On Function and MRNA Expression in Immune Cells in Dairy CowsRoney ZimpelNo ratings yet
- Stress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityDocument18 pagesStress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityCony GSNo ratings yet
- The Ascorbic Acid Transporter SVCT2 Is Expressed in Slow-Twitch Skeletal Muscle WbresDocument10 pagesThe Ascorbic Acid Transporter SVCT2 Is Expressed in Slow-Twitch Skeletal Muscle WbresJorge Eneas DosoliNo ratings yet
- Keystone PosterDocument1 pageKeystone PosterSaswati Chakraborty MisraNo ratings yet
- Menstruation & OvulationDocument42 pagesMenstruation & Ovulationneilesh300No ratings yet
- Renal System - Kristiene JoyDocument11 pagesRenal System - Kristiene JoyijayFLx64No ratings yet
- Alien SpeciesDocument36 pagesAlien SpeciesmkalamarasNo ratings yet
- Research1Document77 pagesResearch1Ces AriaNo ratings yet
- WEEK 8 OTHER BLOOD GROUP SYSTEM PART 1 v2 PDFDocument6 pagesWEEK 8 OTHER BLOOD GROUP SYSTEM PART 1 v2 PDFjohn carlo cabanlitNo ratings yet
- Theories of AgingDocument4 pagesTheories of AgingJClaudz PilapilNo ratings yet
- 2321 FullDocument13 pages2321 FullJuhaydith Hernandez AzuajeNo ratings yet
- Gene Technology Questions AQA OCR EdexcelDocument3 pagesGene Technology Questions AQA OCR EdexcelZainab AlziftawiNo ratings yet
- Identi Fication of Pyridinium With Three Indole Moieties As An Antimicrobial AgentDocument5 pagesIdenti Fication of Pyridinium With Three Indole Moieties As An Antimicrobial AgentLuigi André Cabos ZavaletaNo ratings yet
- Pharmacokinetic and Pharmacodynamic Drug Interactions Associated With Antiretroviral DrugsDocument136 pagesPharmacokinetic and Pharmacodynamic Drug Interactions Associated With Antiretroviral DrugsNenad PanzalovićNo ratings yet
- Psych Unit 4 Study GuideDocument16 pagesPsych Unit 4 Study GuideLauren ThompsonNo ratings yet
- Polymerase Chain Reaction & Types: Nandith P B Research ScholarDocument22 pagesPolymerase Chain Reaction & Types: Nandith P B Research ScholarNandit Banawalikar100% (1)
- MBR 2019 - Biochemistry HandoutsDocument66 pagesMBR 2019 - Biochemistry HandoutsEiaNo ratings yet
- Slide Presentation of Bioethical Issues Concerning Animal Testing in ResearchDocument17 pagesSlide Presentation of Bioethical Issues Concerning Animal Testing in ResearchCt Shahirah100% (1)
- Guidelines RosaceaDocument14 pagesGuidelines Rosaceaika_anggrainiNo ratings yet
- Mula Uddiyan Jalandhar Bandha & Maha MudraDocument4 pagesMula Uddiyan Jalandhar Bandha & Maha MudraBhavesh1801No ratings yet
- Reproductive SystemDocument7 pagesReproductive Systemcollege code - 5No ratings yet
- Unit 5 - Topic 4 PathologyDocument9 pagesUnit 5 - Topic 4 PathologyHùng Mạnh NguyễnNo ratings yet
- BacteriaDocument1 pageBacteriaabdo aliNo ratings yet
- Renal Cell Carcinoma: Diagnosis, Staging, and SurveillanceDocument18 pagesRenal Cell Carcinoma: Diagnosis, Staging, and SurveillanceMutia IsanaNo ratings yet
- Methane Fact SheetDocument6 pagesMethane Fact SheetArshad MahmoodNo ratings yet
- Engineering A New Mouse Model For Vitiligo 2012 Journal of Investigative DermatologyDocument4 pagesEngineering A New Mouse Model For Vitiligo 2012 Journal of Investigative DermatologyAurelian Corneliu MoraruNo ratings yet
- Albumin Based Nanodelivery PDFDocument33 pagesAlbumin Based Nanodelivery PDFFARIS NATTUKALNo ratings yet
- Book Review Why We Get Sick The New Science of DarDocument5 pagesBook Review Why We Get Sick The New Science of DarKhang ChuNo ratings yet
- T Cell Maturation, Activation and DifferentiationDocument65 pagesT Cell Maturation, Activation and DifferentiationChanDan RjNo ratings yet
- Topic Test - Genetics MemoDocument5 pagesTopic Test - Genetics Memomaiselatebogo4No ratings yet
- Jurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootDocument6 pagesJurnal Ilmu Konservasi Gigi: PSA Pada Supernumery RootAchmad Zam Zam AghazyNo ratings yet
- Cell and Cell Division AnswerDocument8 pagesCell and Cell Division AnswerA-0315-12 PRASHARADHA S KUMARNo ratings yet