Download as pdf or txt
You might also like
- Admission NotesDocument16 pagesAdmission NotesCaisar Dewi MaulinaNo ratings yet
- Case Study On Fluid OverloadDocument37 pagesCase Study On Fluid OverloadradicalmpNo ratings yet
- Etiology of MalocclusionDocument185 pagesEtiology of MalocclusionSumita Jain100% (2)
- Pathway Gagal Ginjal Kronik: Intoleransi AktivitasDocument1 pagePathway Gagal Ginjal Kronik: Intoleransi AktivitasReza ApriandiNo ratings yet
- Fu-Ming Zhou and John J. Hablitz - Activation of Serotonin Receptors Modulates Synaptic Transmission in Rat Cerebral CortexDocument12 pagesFu-Ming Zhou and John J. Hablitz - Activation of Serotonin Receptors Modulates Synaptic Transmission in Rat Cerebral CortexCortate15gNo ratings yet
- SCNTFC RepDocument12 pagesSCNTFC RepRehaan FayazNo ratings yet
- Jurnal HomeostatisDocument13 pagesJurnal HomeostatisJastip YueNo ratings yet
- Effects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaDocument12 pagesEffects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaIqbal AbdillahNo ratings yet
- (Sici) 1096 9861 (20000207) 417:2 181::aid Cne4 3.0.co 2 ADocument14 pages(Sici) 1096 9861 (20000207) 417:2 181::aid Cne4 3.0.co 2 AgslifirsNo ratings yet
- HHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDocument22 pagesHHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDebbyNo ratings yet
- 9732 FullDocument10 pages9732 FullDaniel Isaac Castillo HidalgoNo ratings yet
- Li-2020-GABAergic Neurons in The Dorsal Raphe Nucleus That Express 5-HT3A Receptors Participate in Responses To Stress HormonesDocument9 pagesLi-2020-GABAergic Neurons in The Dorsal Raphe Nucleus That Express 5-HT3A Receptors Participate in Responses To Stress HormonesOmar Hernández GonzálezNo ratings yet
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenNo ratings yet
- Corse Et Al. - 1999 - Preclinical Testing of Neuroprotective Neurotrophic Factors in A Model of Chronic Motor Neuron DegenerationDocument12 pagesCorse Et Al. - 1999 - Preclinical Testing of Neuroprotective Neurotrophic Factors in A Model of Chronic Motor Neuron Degenerationmarej312No ratings yet
- Progesterone Synthesis in The Nervous System: Implications For Myelination and Myelin RepairDocument22 pagesProgesterone Synthesis in The Nervous System: Implications For Myelination and Myelin Repair147Anastaysia Anisa HaribaikNo ratings yet
- Molecular and Cellular NeuroscienceDocument8 pagesMolecular and Cellular NeuroscienceDefi DestyawenyNo ratings yet
- Neurobiology of Disease: E.N. Mangano, S. Peters, D. Litteljohn, R. So, C. Bethune, J. Bobyn, M. Clarke, S. HayleyDocument14 pagesNeurobiology of Disease: E.N. Mangano, S. Peters, D. Litteljohn, R. So, C. Bethune, J. Bobyn, M. Clarke, S. HayleyShawn HayleyNo ratings yet
- 17-Estradiol Attenuates Hippocampal Neuronal LossDocument9 pages17-Estradiol Attenuates Hippocampal Neuronal LossrodrigounitedNo ratings yet
- Histamine How It Effects Our Brain and Nervous SystemDocument6 pagesHistamine How It Effects Our Brain and Nervous SystemHerbie KawaihauNo ratings yet
- Dhami Et Al-2019-Journal of Neurochemistry PDFDocument18 pagesDhami Et Al-2019-Journal of Neurochemistry PDFJawad A. KhanNo ratings yet
- Serotonin in Animal Cognition and Behavior: Molecular SciencesDocument23 pagesSerotonin in Animal Cognition and Behavior: Molecular SciencesRifqi Hamdani PasaribuNo ratings yet
- Endo 3955Document7 pagesEndo 3955Isa florNo ratings yet
- Paper 1Document11 pagesPaper 1Anonymous K8fT6HNo ratings yet
- Effects of Psilocybin On Hippocampal Neurogenesis and Extinction of Trace Fear ConditioningDocument11 pagesEffects of Psilocybin On Hippocampal Neurogenesis and Extinction of Trace Fear ConditioningurkogNo ratings yet
- Journal AphasiaDocument9 pagesJournal AphasiaChintya Anugrah SuhendraNo ratings yet
- Iah SDLPDFDocument9 pagesIah SDLPDFMeg LiebNo ratings yet
- The Role of 5-HT Systems on Memory and Dysfunctional Memory: Emergent Targets for Memory Formation and Memory AlterationsFrom EverandThe Role of 5-HT Systems on Memory and Dysfunctional Memory: Emergent Targets for Memory Formation and Memory AlterationsNo ratings yet
- Control of Dorsal Raphe Serotonergic Neurons by The Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABA, and Glutamate ReceptorsDocument13 pagesControl of Dorsal Raphe Serotonergic Neurons by The Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABA, and Glutamate ReceptorsCortate15gNo ratings yet
- Andsberg 1998 EJNDocument11 pagesAndsberg 1998 EJNZaal KokaiaNo ratings yet
- Serotonin and Molecular Neuroimaging in Humans Using PET PDFDocument19 pagesSerotonin and Molecular Neuroimaging in Humans Using PET PDFWynn TheinNo ratings yet
- Serotonin and Beyond: Therapeutics For Major Depression: ReviewDocument7 pagesSerotonin and Beyond: Therapeutics For Major Depression: ReviewmarielaNo ratings yet
- Nutritional FactorsDocument8 pagesNutritional FactorsMagnusAngeltveitKjølenNo ratings yet
- 5-HTT Independent Effects of Fluoxetine On NeuroplasticityDocument11 pages5-HTT Independent Effects of Fluoxetine On NeuroplasticityCony GSNo ratings yet
- Dopamine and Sleep REMDocument13 pagesDopamine and Sleep REMcalfupanNo ratings yet
- 10.1515 - JBCPP 2017 0022Document2 pages10.1515 - JBCPP 2017 0022Hot Parsaulian SiregarNo ratings yet
- Reversing Aging Related DeficitsDocument51 pagesReversing Aging Related DeficitsQuaxi1954No ratings yet
- Ijnp Pyv045 FullDocument35 pagesIjnp Pyv045 FullNityananda PortelladaNo ratings yet
- C3a and HI-neurodegenerationDocument32 pagesC3a and HI-neurodegenerationJingyun WuNo ratings yet
- Perinatal Antidepressant Exposure Alters Cortical Network Function in RodentsDocument6 pagesPerinatal Antidepressant Exposure Alters Cortical Network Function in RodentsAkmal NurulNo ratings yet
- tmp490 TMPDocument11 pagestmp490 TMPFrontiersNo ratings yet
- J Jns 2004 12 011Document10 pagesJ Jns 2004 12 011Viviana LunaNo ratings yet
- Kong Lee Final 1Document10 pagesKong Lee Final 1api-557190611No ratings yet
- Iranian Journal of Basic Medical SciencesDocument10 pagesIranian Journal of Basic Medical SciencesmusmuafeerahNo ratings yet
- Should We Be Anesthetizing Young Children?: Jerrold Lerman MD, FRCPC, FANZCADocument3 pagesShould We Be Anesthetizing Young Children?: Jerrold Lerman MD, FRCPC, FANZCAhugo roblesNo ratings yet
- 1) Yan (2014)Document8 pages1) Yan (2014)Êndel AlvesNo ratings yet
- Role of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyDocument10 pagesRole of Central Vagal 5-HT Receptors in Gastrointestinal Physiology and PathophysiologyStella NoviaNo ratings yet
- Uceyler2010 Lack of The Serotonin Transporter in Mice Reduces Locomotor Activity and Leads To Gender-Dependent Late Onset ObesityDocument11 pagesUceyler2010 Lack of The Serotonin Transporter in Mice Reduces Locomotor Activity and Leads To Gender-Dependent Late Onset ObesityArizonaNo ratings yet
- JNC 162 128Document19 pagesJNC 162 128Jose Alberto SVNo ratings yet
- 2020 - Phosphorylated Tau Interactome in The Human Alzheimer's Disease BrainDocument15 pages2020 - Phosphorylated Tau Interactome in The Human Alzheimer's Disease BrainEl Tal RuleiroNo ratings yet
- Social Isolation Selectively Reduces Hip PDFDocument3 pagesSocial Isolation Selectively Reduces Hip PDFJean Pierre Chastre LuzaNo ratings yet
- Accepted Manuscript: Neurobiology of Learning and MemoryDocument49 pagesAccepted Manuscript: Neurobiology of Learning and MemoryChristine HandayaniNo ratings yet
- Doi10.1016-J.pnpbp.2008.03.024 2011Document8 pagesDoi10.1016-J.pnpbp.2008.03.024 2011Mark BloomanNo ratings yet
- s12035 023 03860 4Document19 pagess12035 023 03860 4Isa florNo ratings yet
- Monique Vallee Et Al - Neurosteroids: Deficient Cognitive Performance in Aged Rats Depends On Low Pregnenolone Sulfate Levels in The HippocampusDocument6 pagesMonique Vallee Et Al - Neurosteroids: Deficient Cognitive Performance in Aged Rats Depends On Low Pregnenolone Sulfate Levels in The HippocampusLonkesNo ratings yet
- Neurobiology of Disease: A B A A A D C A 1 A B D 1Document12 pagesNeurobiology of Disease: A B A A A D C A 1 A B D 1Duxan Arancibia RadichNo ratings yet
- Nhsjs SubmissionDocument8 pagesNhsjs Submissionapi-410186016No ratings yet
- Esearch Rticle: Requirement of Hippocampal Neurogenesis For The Behavioral Effects of AntidepressantsDocument5 pagesEsearch Rticle: Requirement of Hippocampal Neurogenesis For The Behavioral Effects of AntidepressantsMariana JuncoNo ratings yet
- 1a. 24609742 Neurobiology of PainDocument6 pages1a. 24609742 Neurobiology of PainJason HillNo ratings yet
- A9RC92EDocument19 pagesA9RC92ENha MạchNo ratings yet
- Taler 2013Document5 pagesTaler 2013ArouetNo ratings yet
- Brain Lesions, Obesity, and Other Disturbances in Mice Treated With Monosodium Glutamate (Olney 1969)Document3 pagesBrain Lesions, Obesity, and Other Disturbances in Mice Treated With Monosodium Glutamate (Olney 1969)Aprilia OanimaNo ratings yet
- BV2 Publikacja Final Version 06.02.22 For SendingDocument11 pagesBV2 Publikacja Final Version 06.02.22 For SendingEwelina HonkiszNo ratings yet
- Ijpbs 2011Document8 pagesIjpbs 2011Ramakrishna DevakiNo ratings yet
- Thrombin and Brain Recovery After Intracerebral HemorrhageDocument3 pagesThrombin and Brain Recovery After Intracerebral HemorrhageAndhy IndriyonoNo ratings yet
- Nucleosome (1) Nature 2005Document7 pagesNucleosome (1) Nature 2005damien333No ratings yet
- The Two Micron All Sky Survey (2mass) : 2006. The American Astronomical Society. All Rights Reserved. Printed in U.S.ADocument21 pagesThe Two Micron All Sky Survey (2mass) : 2006. The American Astronomical Society. All Rights Reserved. Printed in U.S.Adamien333No ratings yet
- LLBB TrafficDossierDocument23 pagesLLBB TrafficDossierdamien333No ratings yet
- The Sloan Digital Sky Survey: Technical SummaryDocument22 pagesThe Sloan Digital Sky Survey: Technical Summarydamien333No ratings yet
- Guiding Neutral Atoms Around Curves With Lithographically Patterned Current-Carrying WiresDocument9 pagesGuiding Neutral Atoms Around Curves With Lithographically Patterned Current-Carrying Wiresdamien333No ratings yet
- 02 Kohler Keysers Rizzolatti MirrorNeuronsDocument3 pages02 Kohler Keysers Rizzolatti MirrorNeuronsdamien333No ratings yet
- A 1000-Loci Transcript Map of The Barley Genome New Anchoring Points For Integrative Grass GenomicsDocument17 pagesA 1000-Loci Transcript Map of The Barley Genome New Anchoring Points For Integrative Grass Genomicsdamien333No ratings yet
- Sevelamer Attenuates The Progression of Coronary and Aortic Calcification in Hemodialysis PatientsDocument8 pagesSevelamer Attenuates The Progression of Coronary and Aortic Calcification in Hemodialysis Patientsdamien333No ratings yet
- Goldinger Etal JML 97Document27 pagesGoldinger Etal JML 97damien333No ratings yet
- Water and Salinity Stress in Grapevines: Early and Late Changes in Transcript and Metabolite ProfilesDocument24 pagesWater and Salinity Stress in Grapevines: Early and Late Changes in Transcript and Metabolite Profilesdamien333No ratings yet
- Comparative Genomics of The HOG-signalling System in Fungi: Ó Springer-Verlag 2006Document15 pagesComparative Genomics of The HOG-signalling System in Fungi: Ó Springer-Verlag 2006damien333No ratings yet
- Dynamics of The Lake Michigan Food Web1970-2000Document18 pagesDynamics of The Lake Michigan Food Web1970-2000damien333No ratings yet
- Magnetic Field0602194Document4 pagesMagnetic Field0602194damien333No ratings yet
- Web Question Answering: Is More Always Better?: Susan Dumais, Michele Banko, Eric Brill, Jimmy Lin, Andrew NGDocument9 pagesWeb Question Answering: Is More Always Better?: Susan Dumais, Michele Banko, Eric Brill, Jimmy Lin, Andrew NGdamien333No ratings yet
- Chest Pain - Part 1 - Initial Presentation & InvestigationsDocument4 pagesChest Pain - Part 1 - Initial Presentation & InvestigationsThorgan HazardNo ratings yet
- Free Radical 2Document20 pagesFree Radical 2Munewer AbdellaNo ratings yet
- Hydrocephalus PPT SSMC RewaDocument60 pagesHydrocephalus PPT SSMC RewaAbhishek Mishra100% (3)
- Acute Rheumatic Fever PathophysiologyDocument1 pageAcute Rheumatic Fever PathophysiologyMoonyeen Jann Casera BalicNo ratings yet
- NIFEDIPINE Extended Release Tablets 30 MG, 60 MG, and 90 MGDocument13 pagesNIFEDIPINE Extended Release Tablets 30 MG, 60 MG, and 90 MGXICMENNo ratings yet
- Chapter11 PDFDocument24 pagesChapter11 PDFsgw67No ratings yet
- Brain Arteriovenous Malforamtion: Dr. Suresh Bishokarma MS, MCH (Neurosurgery)Document95 pagesBrain Arteriovenous Malforamtion: Dr. Suresh Bishokarma MS, MCH (Neurosurgery)rr_eeyNo ratings yet
- Panvascular Disease - Diagnosis and Management: SciencedirectDocument9 pagesPanvascular Disease - Diagnosis and Management: SciencedirectAndikaputra Brahma WidiantoroNo ratings yet
- Paediatric Hospital Level Stgs and Eml - 4th Ed 2017 - Final-Ilovepdf-CompressedDocument656 pagesPaediatric Hospital Level Stgs and Eml - 4th Ed 2017 - Final-Ilovepdf-CompressedMeshaki MbarukaNo ratings yet
- Gangguan Asam BasaDocument51 pagesGangguan Asam BasaYudhistira YuliandraNo ratings yet
- Osms - It/alzheimers-Disease: Pathology & CausesDocument1 pageOsms - It/alzheimers-Disease: Pathology & Causesdysa ayu shalsabilaNo ratings yet
- NorgestrelDocument3 pagesNorgestrelapi-3797941No ratings yet
- (Ebook pdf) Pharmacotherapy Casebook: A Patient-Focused Approach, 11th Edition Terry L. Schwinghammer - eBook PDF all chapterDocument69 pages(Ebook pdf) Pharmacotherapy Casebook: A Patient-Focused Approach, 11th Edition Terry L. Schwinghammer - eBook PDF all chapterdkairirinej18100% (3)
- OSCE of Ear Nose & ThroatDocument23 pagesOSCE of Ear Nose & Throatmasood alam100% (1)
- Oedematous Children OSCE Nephrotic NephriticDocument5 pagesOedematous Children OSCE Nephrotic NephriticJJ LimNo ratings yet
- PONSTAN® Capsules: What Is in This LeafletDocument4 pagesPONSTAN® Capsules: What Is in This LeafletLee Hye RiNo ratings yet
- Clinical Questions PICO and Study Design EBM Part 1Document47 pagesClinical Questions PICO and Study Design EBM Part 1Yaser ZaherNo ratings yet
- Alzheimer's Disease: General InformationsDocument2 pagesAlzheimer's Disease: General InformationsRausz KatalinNo ratings yet
- Hemorrhagic Shock Classification in Pediatric TraumaDocument1 pageHemorrhagic Shock Classification in Pediatric TraumaPrisia AnantamaNo ratings yet
- Bulbar and Pseudobulbar Palsy: Search LITFL ..Document8 pagesBulbar and Pseudobulbar Palsy: Search LITFL ..Utkarsh ShahiNo ratings yet
- 132 Emergency MedicineDocument14 pages132 Emergency MedicineVania NandaNo ratings yet
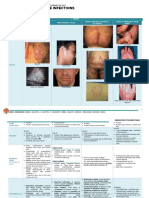
- (DERMA) 03 TineasDocument9 pages(DERMA) 03 TineasJolaine ValloNo ratings yet
- Sheena The EyeDocument9 pagesSheena The EyeSheena Yen de Pano-PagdalianNo ratings yet
- Armour Thyroid (Thyroid Tablets, USP) RX Only: I I NHDocument10 pagesArmour Thyroid (Thyroid Tablets, USP) RX Only: I I NHAdam GolsbyNo ratings yet
- PediatricsDocument21 pagesPediatricsJo Anne75% (8)
- Prostate CancerDocument67 pagesProstate CancerMalueth AnguiNo ratings yet