
Plants and Inorganic Nutrients: The Essential Nutrient Elements
Plants and Inorganic Nutrients: The Essential Nutrient Elements
You might also like
- Moles PacketDocument11 pagesMoles PacketCaslyn Campbell0% (1)
- KIZEN - Infinite Off SeasonDocument13 pagesKIZEN - Infinite Off SeasonLeon50% (4)
- Ebin Belderol TB and WorksheetDocument11 pagesEbin Belderol TB and WorksheetMarielle Ebin100% (3)
- ASTM 6365 - 99 - Spark TestDocument4 pagesASTM 6365 - 99 - Spark Testjudith_ayala_10No ratings yet
- Conmed Sabre 180Document54 pagesConmed Sabre 180ClaudiaNavarreteNo ratings yet
- Plant NutrientsDocument10 pagesPlant NutrientsAdrian GligaNo ratings yet
- Mineral Nutrition: Joanna Marie Betito Mile - BiologyDocument30 pagesMineral Nutrition: Joanna Marie Betito Mile - BiologyEricka MarieNo ratings yet
- Lesson 3 Plant and Animal Organ Systems and Their Functions (Nutrition)Document11 pagesLesson 3 Plant and Animal Organ Systems and Their Functions (Nutrition)Ishs GwsNo ratings yet
- Las 3 Genbio2Document15 pagesLas 3 Genbio2hellohelloworldNo ratings yet
- Lec05jurnal AuksinDocument14 pagesLec05jurnal AuksinMoch Rosyadi AdnanNo ratings yet
- C. Let's Check: Pacalna, Anifah B. BSA-3 Year Week 1 UloaDocument17 pagesC. Let's Check: Pacalna, Anifah B. BSA-3 Year Week 1 UloaJunzen Ralph YapNo ratings yet
- Mineral NutritionDocument23 pagesMineral NutritionRahul KumarNo ratings yet
- Lemna SP ArmDocument16 pagesLemna SP ArmHanin Jaliyy100% (1)
- Soil Biology Primer by Elaine R. Ingham-with-cover-page-V2Document22 pagesSoil Biology Primer by Elaine R. Ingham-with-cover-page-V2vincentNo ratings yet
- Plant Nutrition Is The Study of The Chemical Elements That Are Necessary For GrowthDocument24 pagesPlant Nutrition Is The Study of The Chemical Elements That Are Necessary For GrowthDianna Ipray GriffithsNo ratings yet
- NUTRITIONDocument26 pagesNUTRITIONchastine daneNo ratings yet
- Ishrana Biljaka U Rasadnicima - PredavanjaDocument14 pagesIshrana Biljaka U Rasadnicima - PredavanjaDanijel JurisicNo ratings yet
- Soil Biology The Soil Biology Primer ChaDocument21 pagesSoil Biology The Soil Biology Primer ChaRojotiana Marco RAKOTOMANANANo ratings yet
- Nutrition in Plants NotesDocument7 pagesNutrition in Plants NotesMidhun Bhuvanesh.B 7ANo ratings yet
- Plant Nutrition - WikipediaDocument87 pagesPlant Nutrition - WikipediaBashiir NuurNo ratings yet
- Fertilizer For TomatoesDocument8 pagesFertilizer For TomatoesIsabel Sofia Corredor MendezNo ratings yet
- New Microsoft Word DocumentDocument12 pagesNew Microsoft Word DocumentAkash BhardwajNo ratings yet
- NutritionDocument10 pagesNutritionJerwin GarnaceNo ratings yet
- Nutrition in Plants Class 7 Science Notes - Chapter 1Document5 pagesNutrition in Plants Class 7 Science Notes - Chapter 1Yash ArdeshnaNo ratings yet
- Rhizospheric & PhyllophericDocument6 pagesRhizospheric & PhyllophericDibyakNo ratings yet
- DavongDocument4 pagesDavongArthadian De PeraltaNo ratings yet
- Chapter 37 Plant Nutrition Outline: A Nutritional NetworkDocument15 pagesChapter 37 Plant Nutrition Outline: A Nutritional Networkmario hasanahNo ratings yet
- Nhóm 1 - Phosphate Solubilizing Bacteria and Fungi From Rhizosphere of PlantsDocument27 pagesNhóm 1 - Phosphate Solubilizing Bacteria and Fungi From Rhizosphere of Plantsthanhhnga1623No ratings yet
- © Ncert Not To Be Republished: Nutrition in PlantsDocument10 pages© Ncert Not To Be Republished: Nutrition in PlantsKalpavriksha1974No ratings yet
- Plant Mineral Nutrition 2014 FinalpdfDocument7 pagesPlant Mineral Nutrition 2014 Finalpdfalhajifawaz77No ratings yet
- Lecture 2B Plant NutrientDocument39 pagesLecture 2B Plant NutrientFarah ReginaNo ratings yet
- Course Name-Biochemistry and Plant Physiology (Paper Code-BOT 504)Document24 pagesCourse Name-Biochemistry and Plant Physiology (Paper Code-BOT 504)selularbintangmasNo ratings yet
- Reading and Listening Vol. 1 Test 2.Document44 pagesReading and Listening Vol. 1 Test 2.shengchihsuNo ratings yet
- Lesson 3 PLANT AND ANIMAL NUTRITIONDocument6 pagesLesson 3 PLANT AND ANIMAL NUTRITIONNicoleNo ratings yet
- 22 Introduction To Foliar Fertilizers HS - Sergey PigarevDocument21 pages22 Introduction To Foliar Fertilizers HS - Sergey PigarevGhada HindawyNo ratings yet
- TodayDocument5 pagesTodayABEBE AMARENo ratings yet
- Review Article: Antioxidant Functions of Phytic AcidDocument9 pagesReview Article: Antioxidant Functions of Phytic AcidPhạm Nguyễn Minh ThuNo ratings yet
- The Nitrogen CycleDocument19 pagesThe Nitrogen CycleLiam Nathaniel V. SolisNo ratings yet
- Chapter 5-7Document38 pagesChapter 5-7mekonnenNo ratings yet
- Nutrition (XI) in PlantsDocument56 pagesNutrition (XI) in PlantsMehak MuradaniNo ratings yet
- CH 27Document2 pagesCH 27Austin JungNo ratings yet
- Root Nodules (Rhizobium Legumes) PDFDocument2 pagesRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07No ratings yet
- Membrane UnitDocument15 pagesMembrane UnitHalo BashirNo ratings yet
- Module I - Soil 2Document6 pagesModule I - Soil 2cristine abridaNo ratings yet
- Nutrition Presentation Doc NDocument4 pagesNutrition Presentation Doc NSumaira BibiNo ratings yet
- Plant Nutrition NotesDocument4 pagesPlant Nutrition Notesvijend100% (1)
- Plant Tissue Culture MediaDocument17 pagesPlant Tissue Culture MediaNorhisham DaudNo ratings yet
- The Soil Food Web PDFDocument5 pagesThe Soil Food Web PDFAhmed Al-AlawiNo ratings yet
- Predators and Their Prey: Herbivore Predation On PlantsDocument9 pagesPredators and Their Prey: Herbivore Predation On PlantshomamunfatNo ratings yet
- Trabajo Grapal de Ingles ActualDocument4 pagesTrabajo Grapal de Ingles ActualVini MarcosNo ratings yet
- Fungal EcologyDocument21 pagesFungal EcologyLiselle Pillay100% (1)
- Nutrition in Plants Class 7 Science NotesDocument4 pagesNutrition in Plants Class 7 Science Notesshravandownload100% (1)
- Nutrition in PlantsDocument44 pagesNutrition in Plantsgeorgy shibuNo ratings yet
- Micronutrients: They Are Also Called Trace Elements and Are Present in Plant Bodies in Very Small AmountsDocument4 pagesMicronutrients: They Are Also Called Trace Elements and Are Present in Plant Bodies in Very Small AmountsNittu MaryNo ratings yet
- Lemna Is A Genus of Free-FloatingDocument20 pagesLemna Is A Genus of Free-FloatingnsalmiyahNo ratings yet
- Delhi Public School - Bopal, Ahmedabad (2021-22) : Class: VII Subject: Science Chapter 1 Nutrition in Plants Study NotesDocument10 pagesDelhi Public School - Bopal, Ahmedabad (2021-22) : Class: VII Subject: Science Chapter 1 Nutrition in Plants Study Notesbhargav guptaNo ratings yet
- The Soil Food Web: Tuning in To The World Beneath Our FeetDocument5 pagesThe Soil Food Web: Tuning in To The World Beneath Our Feethainy09No ratings yet
- 472 - Bio 103 WeboDocument8 pages472 - Bio 103 Weboabiolaelizabeth255No ratings yet
- STPM Biology Chapter 6 NotesDocument8 pagesSTPM Biology Chapter 6 NotesVentus Tan100% (1)
- Plant Mineral Nutrition March 2024Document16 pagesPlant Mineral Nutrition March 2024Paramveer SinghNo ratings yet
- Handbook of Fertilizers - Their Sources, Make-Up, Effects, and UseFrom EverandHandbook of Fertilizers - Their Sources, Make-Up, Effects, and UseRating: 4.5 out of 5 stars4.5/5 (4)
- KSEB Online Payment - Transaction DetailsDocument1 pageKSEB Online Payment - Transaction DetailsAsish GeiorgeNo ratings yet
- Case Report: Nasal Myiasis in An Elderly Patient With Atrophic Rhinitis and Facial Sequelae of LeprosyDocument3 pagesCase Report: Nasal Myiasis in An Elderly Patient With Atrophic Rhinitis and Facial Sequelae of LeprosyAsish GeiorgeNo ratings yet
- English Specimen Paper 1 2018 PDFDocument8 pagesEnglish Specimen Paper 1 2018 PDFAsish GeiorgeNo ratings yet
- The Oesophagus, Stomach and Small Bowel: TrunkDocument11 pagesThe Oesophagus, Stomach and Small Bowel: TrunkAsish GeiorgeNo ratings yet
- The Rectum and Anus: TrunkDocument11 pagesThe Rectum and Anus: TrunkAsish GeiorgeNo ratings yet
- Fever With Rash (Peds)Document13 pagesFever With Rash (Peds)Asish Geiorge100% (1)
- The Spleen: Splenic Trauma and Splenectomy: TrunkDocument3 pagesThe Spleen: Splenic Trauma and Splenectomy: TrunkAsish GeiorgeNo ratings yet
- MSC CVS Gottingen SyllabusDocument14 pagesMSC CVS Gottingen SyllabusAsish GeiorgeNo ratings yet
- Nephritis Fact SheetDocument3 pagesNephritis Fact SheetAsish GeiorgeNo ratings yet
- Opthal LO PDFDocument142 pagesOpthal LO PDFAsish GeiorgeNo ratings yet
- The Major Pathways of Complement ActivationDocument2 pagesThe Major Pathways of Complement ActivationAsish GeiorgeNo ratings yet
- FeverDocument45 pagesFeverAsish Geiorge100% (1)
- MSC PHD EndoMetabolismDocument2 pagesMSC PHD EndoMetabolismAsish GeiorgeNo ratings yet
- Estimation of B Cell MassDocument5 pagesEstimation of B Cell MassAsish GeiorgeNo ratings yet
- World's Largest Science, Technology & Medicine Open Access Book PublisherDocument22 pagesWorld's Largest Science, Technology & Medicine Open Access Book PublisherAsish GeiorgeNo ratings yet
- Physiology of Cornea: Corneal TransparencyDocument2 pagesPhysiology of Cornea: Corneal TransparencyAsish GeiorgeNo ratings yet
- 2-BP-3.5K-5K 220v ManualDocument42 pages2-BP-3.5K-5K 220v ManualEcosenseTechnologies GreenenergyNo ratings yet
- Module 2 Project Gap Analysis Strategy For Team #3Document20 pagesModule 2 Project Gap Analysis Strategy For Team #3Mary A. Dillard100% (2)
- Attorney General Report On NM Prison Riot - Part 1Document150 pagesAttorney General Report On NM Prison Riot - Part 1Albuquerque Journal100% (3)
- Clinical Liver Disease - 2022 - Memel - Intermittent Fasting As A Treatment For Nonalcoholic Fatty Liver Disease What IsDocument5 pagesClinical Liver Disease - 2022 - Memel - Intermittent Fasting As A Treatment For Nonalcoholic Fatty Liver Disease What Isarranguezjr5991No ratings yet
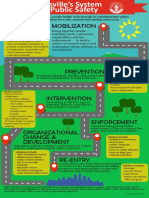
- Reimagining Public SafetyDocument1 pageReimagining Public SafetyWAVE 3 NewsNo ratings yet
- Cells Teachers Guide Discovery EducationDocument38 pagesCells Teachers Guide Discovery EducationKari Kristine Hoskins BarreraNo ratings yet
- Review of Agriculture Robotics: Practicality and FeasibilityDocument6 pagesReview of Agriculture Robotics: Practicality and Feasibilityaldy faleraNo ratings yet
- Transverse Lie : Predisposing Factors, Maternal and Perinatal OutcomeDocument4 pagesTransverse Lie : Predisposing Factors, Maternal and Perinatal OutcomeErlangga DayudNo ratings yet
- Basic Facts About ESOPs2Document20 pagesBasic Facts About ESOPs2Quant TradingNo ratings yet
- Hemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareDocument13 pagesHemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareRandee BoiiNo ratings yet
- Three Main Parts of An Essay: An Introductory Paragraph A Body A Concluding ParagraphDocument10 pagesThree Main Parts of An Essay: An Introductory Paragraph A Body A Concluding ParagraphLuluk NafisahNo ratings yet
- (U) Daily Activity Report: Marshall DistrictDocument5 pages(U) Daily Activity Report: Marshall DistrictFauquier NowNo ratings yet
- CompressorDocument33 pagesCompressorfvaefaNo ratings yet
- Joy Trust Fear Surprise Sadness Disgust Anger AnticipationDocument4 pagesJoy Trust Fear Surprise Sadness Disgust Anger AnticipationsatyakaamsNo ratings yet
- DSFSDFSDF Advanced Construction TechnologiesDocument3 pagesDSFSDFSDF Advanced Construction Technologiessarveshfdk48No ratings yet
- Fluor CarbonDocument3 pagesFluor CarbonHrishikesh DhawadshikarNo ratings yet
- Copper (RJ-45) SFP: Technical SpecificationsDocument1 pageCopper (RJ-45) SFP: Technical SpecificationsDanielLopezNo ratings yet
- Science Journalism ReviewerDocument15 pagesScience Journalism ReviewerHiyami ChanNo ratings yet
- Peugeot 607 Owners Manual 2005Document206 pagesPeugeot 607 Owners Manual 2005AndreiNo ratings yet
- Activity No. 2 Disaster RiskDocument3 pagesActivity No. 2 Disaster RiskBUENAVENTURA, GABRIEL JOVANNo ratings yet
- MYCOSESDocument41 pagesMYCOSESDayana PrasanthNo ratings yet
- The Internet Addiction and The Academic Productivity of Grade 12 Students in Aclc OrmocDocument43 pagesThe Internet Addiction and The Academic Productivity of Grade 12 Students in Aclc OrmocKenberly DingleNo ratings yet
- Jetlax's CNS Pharmacology Cheat Sheet For The Philippines v6.0 at Bit - Ly - CNSPcol - See Bit - Ly - CNSHandouts For 5.0 CorrectionsDocument61 pagesJetlax's CNS Pharmacology Cheat Sheet For The Philippines v6.0 at Bit - Ly - CNSPcol - See Bit - Ly - CNSHandouts For 5.0 CorrectionsShalimar BasmanNo ratings yet
- Pre-Test in Science 6: M.S. Garcia Elementary SchoolDocument3 pagesPre-Test in Science 6: M.S. Garcia Elementary SchoolAnepsu HohoNo ratings yet
- Technical Data Sheet TGZ190 Rev-1.pdf (Read Only)Document2 pagesTechnical Data Sheet TGZ190 Rev-1.pdf (Read Only)Fatiga jrNo ratings yet









































































































Download as doc, pdf, or txt
You might also like
- Moles PacketDocument11 pagesMoles PacketCaslyn Campbell0% (1)
- KIZEN - Infinite Off SeasonDocument13 pagesKIZEN - Infinite Off SeasonLeon50% (4)
- Ebin Belderol TB and WorksheetDocument11 pagesEbin Belderol TB and WorksheetMarielle Ebin100% (3)
- ASTM 6365 - 99 - Spark TestDocument4 pagesASTM 6365 - 99 - Spark Testjudith_ayala_10No ratings yet
- Conmed Sabre 180Document54 pagesConmed Sabre 180ClaudiaNavarreteNo ratings yet
- Plant NutrientsDocument10 pagesPlant NutrientsAdrian GligaNo ratings yet
- Mineral Nutrition: Joanna Marie Betito Mile - BiologyDocument30 pagesMineral Nutrition: Joanna Marie Betito Mile - BiologyEricka MarieNo ratings yet
- Lesson 3 Plant and Animal Organ Systems and Their Functions (Nutrition)Document11 pagesLesson 3 Plant and Animal Organ Systems and Their Functions (Nutrition)Ishs GwsNo ratings yet
- Las 3 Genbio2Document15 pagesLas 3 Genbio2hellohelloworldNo ratings yet
- Lec05jurnal AuksinDocument14 pagesLec05jurnal AuksinMoch Rosyadi AdnanNo ratings yet
- C. Let's Check: Pacalna, Anifah B. BSA-3 Year Week 1 UloaDocument17 pagesC. Let's Check: Pacalna, Anifah B. BSA-3 Year Week 1 UloaJunzen Ralph YapNo ratings yet
- Mineral NutritionDocument23 pagesMineral NutritionRahul KumarNo ratings yet
- Lemna SP ArmDocument16 pagesLemna SP ArmHanin Jaliyy100% (1)
- Soil Biology Primer by Elaine R. Ingham-with-cover-page-V2Document22 pagesSoil Biology Primer by Elaine R. Ingham-with-cover-page-V2vincentNo ratings yet
- Plant Nutrition Is The Study of The Chemical Elements That Are Necessary For GrowthDocument24 pagesPlant Nutrition Is The Study of The Chemical Elements That Are Necessary For GrowthDianna Ipray GriffithsNo ratings yet
- NUTRITIONDocument26 pagesNUTRITIONchastine daneNo ratings yet
- Ishrana Biljaka U Rasadnicima - PredavanjaDocument14 pagesIshrana Biljaka U Rasadnicima - PredavanjaDanijel JurisicNo ratings yet
- Soil Biology The Soil Biology Primer ChaDocument21 pagesSoil Biology The Soil Biology Primer ChaRojotiana Marco RAKOTOMANANANo ratings yet
- Nutrition in Plants NotesDocument7 pagesNutrition in Plants NotesMidhun Bhuvanesh.B 7ANo ratings yet
- Plant Nutrition - WikipediaDocument87 pagesPlant Nutrition - WikipediaBashiir NuurNo ratings yet
- Fertilizer For TomatoesDocument8 pagesFertilizer For TomatoesIsabel Sofia Corredor MendezNo ratings yet
- New Microsoft Word DocumentDocument12 pagesNew Microsoft Word DocumentAkash BhardwajNo ratings yet
- NutritionDocument10 pagesNutritionJerwin GarnaceNo ratings yet
- Nutrition in Plants Class 7 Science Notes - Chapter 1Document5 pagesNutrition in Plants Class 7 Science Notes - Chapter 1Yash ArdeshnaNo ratings yet
- Rhizospheric & PhyllophericDocument6 pagesRhizospheric & PhyllophericDibyakNo ratings yet
- DavongDocument4 pagesDavongArthadian De PeraltaNo ratings yet
- Chapter 37 Plant Nutrition Outline: A Nutritional NetworkDocument15 pagesChapter 37 Plant Nutrition Outline: A Nutritional Networkmario hasanahNo ratings yet
- Nhóm 1 - Phosphate Solubilizing Bacteria and Fungi From Rhizosphere of PlantsDocument27 pagesNhóm 1 - Phosphate Solubilizing Bacteria and Fungi From Rhizosphere of Plantsthanhhnga1623No ratings yet
- © Ncert Not To Be Republished: Nutrition in PlantsDocument10 pages© Ncert Not To Be Republished: Nutrition in PlantsKalpavriksha1974No ratings yet
- Plant Mineral Nutrition 2014 FinalpdfDocument7 pagesPlant Mineral Nutrition 2014 Finalpdfalhajifawaz77No ratings yet
- Lecture 2B Plant NutrientDocument39 pagesLecture 2B Plant NutrientFarah ReginaNo ratings yet
- Course Name-Biochemistry and Plant Physiology (Paper Code-BOT 504)Document24 pagesCourse Name-Biochemistry and Plant Physiology (Paper Code-BOT 504)selularbintangmasNo ratings yet
- Reading and Listening Vol. 1 Test 2.Document44 pagesReading and Listening Vol. 1 Test 2.shengchihsuNo ratings yet
- Lesson 3 PLANT AND ANIMAL NUTRITIONDocument6 pagesLesson 3 PLANT AND ANIMAL NUTRITIONNicoleNo ratings yet
- 22 Introduction To Foliar Fertilizers HS - Sergey PigarevDocument21 pages22 Introduction To Foliar Fertilizers HS - Sergey PigarevGhada HindawyNo ratings yet
- TodayDocument5 pagesTodayABEBE AMARENo ratings yet
- Review Article: Antioxidant Functions of Phytic AcidDocument9 pagesReview Article: Antioxidant Functions of Phytic AcidPhạm Nguyễn Minh ThuNo ratings yet
- The Nitrogen CycleDocument19 pagesThe Nitrogen CycleLiam Nathaniel V. SolisNo ratings yet
- Chapter 5-7Document38 pagesChapter 5-7mekonnenNo ratings yet
- Nutrition (XI) in PlantsDocument56 pagesNutrition (XI) in PlantsMehak MuradaniNo ratings yet
- CH 27Document2 pagesCH 27Austin JungNo ratings yet
- Root Nodules (Rhizobium Legumes) PDFDocument2 pagesRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07No ratings yet
- Membrane UnitDocument15 pagesMembrane UnitHalo BashirNo ratings yet
- Module I - Soil 2Document6 pagesModule I - Soil 2cristine abridaNo ratings yet
- Nutrition Presentation Doc NDocument4 pagesNutrition Presentation Doc NSumaira BibiNo ratings yet
- Plant Nutrition NotesDocument4 pagesPlant Nutrition Notesvijend100% (1)
- Plant Tissue Culture MediaDocument17 pagesPlant Tissue Culture MediaNorhisham DaudNo ratings yet
- The Soil Food Web PDFDocument5 pagesThe Soil Food Web PDFAhmed Al-AlawiNo ratings yet
- Predators and Their Prey: Herbivore Predation On PlantsDocument9 pagesPredators and Their Prey: Herbivore Predation On PlantshomamunfatNo ratings yet
- Trabajo Grapal de Ingles ActualDocument4 pagesTrabajo Grapal de Ingles ActualVini MarcosNo ratings yet
- Fungal EcologyDocument21 pagesFungal EcologyLiselle Pillay100% (1)
- Nutrition in Plants Class 7 Science NotesDocument4 pagesNutrition in Plants Class 7 Science Notesshravandownload100% (1)
- Nutrition in PlantsDocument44 pagesNutrition in Plantsgeorgy shibuNo ratings yet
- Micronutrients: They Are Also Called Trace Elements and Are Present in Plant Bodies in Very Small AmountsDocument4 pagesMicronutrients: They Are Also Called Trace Elements and Are Present in Plant Bodies in Very Small AmountsNittu MaryNo ratings yet
- Lemna Is A Genus of Free-FloatingDocument20 pagesLemna Is A Genus of Free-FloatingnsalmiyahNo ratings yet
- Delhi Public School - Bopal, Ahmedabad (2021-22) : Class: VII Subject: Science Chapter 1 Nutrition in Plants Study NotesDocument10 pagesDelhi Public School - Bopal, Ahmedabad (2021-22) : Class: VII Subject: Science Chapter 1 Nutrition in Plants Study Notesbhargav guptaNo ratings yet
- The Soil Food Web: Tuning in To The World Beneath Our FeetDocument5 pagesThe Soil Food Web: Tuning in To The World Beneath Our Feethainy09No ratings yet
- 472 - Bio 103 WeboDocument8 pages472 - Bio 103 Weboabiolaelizabeth255No ratings yet
- STPM Biology Chapter 6 NotesDocument8 pagesSTPM Biology Chapter 6 NotesVentus Tan100% (1)
- Plant Mineral Nutrition March 2024Document16 pagesPlant Mineral Nutrition March 2024Paramveer SinghNo ratings yet
- Handbook of Fertilizers - Their Sources, Make-Up, Effects, and UseFrom EverandHandbook of Fertilizers - Their Sources, Make-Up, Effects, and UseRating: 4.5 out of 5 stars4.5/5 (4)
- KSEB Online Payment - Transaction DetailsDocument1 pageKSEB Online Payment - Transaction DetailsAsish GeiorgeNo ratings yet
- Case Report: Nasal Myiasis in An Elderly Patient With Atrophic Rhinitis and Facial Sequelae of LeprosyDocument3 pagesCase Report: Nasal Myiasis in An Elderly Patient With Atrophic Rhinitis and Facial Sequelae of LeprosyAsish GeiorgeNo ratings yet
- English Specimen Paper 1 2018 PDFDocument8 pagesEnglish Specimen Paper 1 2018 PDFAsish GeiorgeNo ratings yet
- The Oesophagus, Stomach and Small Bowel: TrunkDocument11 pagesThe Oesophagus, Stomach and Small Bowel: TrunkAsish GeiorgeNo ratings yet
- The Rectum and Anus: TrunkDocument11 pagesThe Rectum and Anus: TrunkAsish GeiorgeNo ratings yet
- Fever With Rash (Peds)Document13 pagesFever With Rash (Peds)Asish Geiorge100% (1)
- The Spleen: Splenic Trauma and Splenectomy: TrunkDocument3 pagesThe Spleen: Splenic Trauma and Splenectomy: TrunkAsish GeiorgeNo ratings yet
- MSC CVS Gottingen SyllabusDocument14 pagesMSC CVS Gottingen SyllabusAsish GeiorgeNo ratings yet
- Nephritis Fact SheetDocument3 pagesNephritis Fact SheetAsish GeiorgeNo ratings yet
- Opthal LO PDFDocument142 pagesOpthal LO PDFAsish GeiorgeNo ratings yet
- The Major Pathways of Complement ActivationDocument2 pagesThe Major Pathways of Complement ActivationAsish GeiorgeNo ratings yet
- FeverDocument45 pagesFeverAsish Geiorge100% (1)
- MSC PHD EndoMetabolismDocument2 pagesMSC PHD EndoMetabolismAsish GeiorgeNo ratings yet
- Estimation of B Cell MassDocument5 pagesEstimation of B Cell MassAsish GeiorgeNo ratings yet
- World's Largest Science, Technology & Medicine Open Access Book PublisherDocument22 pagesWorld's Largest Science, Technology & Medicine Open Access Book PublisherAsish GeiorgeNo ratings yet
- Physiology of Cornea: Corneal TransparencyDocument2 pagesPhysiology of Cornea: Corneal TransparencyAsish GeiorgeNo ratings yet
- 2-BP-3.5K-5K 220v ManualDocument42 pages2-BP-3.5K-5K 220v ManualEcosenseTechnologies GreenenergyNo ratings yet
- Module 2 Project Gap Analysis Strategy For Team #3Document20 pagesModule 2 Project Gap Analysis Strategy For Team #3Mary A. Dillard100% (2)
- Attorney General Report On NM Prison Riot - Part 1Document150 pagesAttorney General Report On NM Prison Riot - Part 1Albuquerque Journal100% (3)
- Clinical Liver Disease - 2022 - Memel - Intermittent Fasting As A Treatment For Nonalcoholic Fatty Liver Disease What IsDocument5 pagesClinical Liver Disease - 2022 - Memel - Intermittent Fasting As A Treatment For Nonalcoholic Fatty Liver Disease What Isarranguezjr5991No ratings yet
- Reimagining Public SafetyDocument1 pageReimagining Public SafetyWAVE 3 NewsNo ratings yet
- Cells Teachers Guide Discovery EducationDocument38 pagesCells Teachers Guide Discovery EducationKari Kristine Hoskins BarreraNo ratings yet
- Review of Agriculture Robotics: Practicality and FeasibilityDocument6 pagesReview of Agriculture Robotics: Practicality and Feasibilityaldy faleraNo ratings yet
- Transverse Lie : Predisposing Factors, Maternal and Perinatal OutcomeDocument4 pagesTransverse Lie : Predisposing Factors, Maternal and Perinatal OutcomeErlangga DayudNo ratings yet
- Basic Facts About ESOPs2Document20 pagesBasic Facts About ESOPs2Quant TradingNo ratings yet
- Hemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareDocument13 pagesHemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareRandee BoiiNo ratings yet
- Three Main Parts of An Essay: An Introductory Paragraph A Body A Concluding ParagraphDocument10 pagesThree Main Parts of An Essay: An Introductory Paragraph A Body A Concluding ParagraphLuluk NafisahNo ratings yet
- (U) Daily Activity Report: Marshall DistrictDocument5 pages(U) Daily Activity Report: Marshall DistrictFauquier NowNo ratings yet
- CompressorDocument33 pagesCompressorfvaefaNo ratings yet
- Joy Trust Fear Surprise Sadness Disgust Anger AnticipationDocument4 pagesJoy Trust Fear Surprise Sadness Disgust Anger AnticipationsatyakaamsNo ratings yet
- DSFSDFSDF Advanced Construction TechnologiesDocument3 pagesDSFSDFSDF Advanced Construction Technologiessarveshfdk48No ratings yet
- Fluor CarbonDocument3 pagesFluor CarbonHrishikesh DhawadshikarNo ratings yet
- Copper (RJ-45) SFP: Technical SpecificationsDocument1 pageCopper (RJ-45) SFP: Technical SpecificationsDanielLopezNo ratings yet
- Science Journalism ReviewerDocument15 pagesScience Journalism ReviewerHiyami ChanNo ratings yet
- Peugeot 607 Owners Manual 2005Document206 pagesPeugeot 607 Owners Manual 2005AndreiNo ratings yet
- Activity No. 2 Disaster RiskDocument3 pagesActivity No. 2 Disaster RiskBUENAVENTURA, GABRIEL JOVANNo ratings yet
- MYCOSESDocument41 pagesMYCOSESDayana PrasanthNo ratings yet
- The Internet Addiction and The Academic Productivity of Grade 12 Students in Aclc OrmocDocument43 pagesThe Internet Addiction and The Academic Productivity of Grade 12 Students in Aclc OrmocKenberly DingleNo ratings yet
- Jetlax's CNS Pharmacology Cheat Sheet For The Philippines v6.0 at Bit - Ly - CNSPcol - See Bit - Ly - CNSHandouts For 5.0 CorrectionsDocument61 pagesJetlax's CNS Pharmacology Cheat Sheet For The Philippines v6.0 at Bit - Ly - CNSPcol - See Bit - Ly - CNSHandouts For 5.0 CorrectionsShalimar BasmanNo ratings yet
- Pre-Test in Science 6: M.S. Garcia Elementary SchoolDocument3 pagesPre-Test in Science 6: M.S. Garcia Elementary SchoolAnepsu HohoNo ratings yet
- Technical Data Sheet TGZ190 Rev-1.pdf (Read Only)Document2 pagesTechnical Data Sheet TGZ190 Rev-1.pdf (Read Only)Fatiga jrNo ratings yet