Download as pdf or txt
You might also like
- SECTION 1 Tim TurnerDocument61 pagesSECTION 1 Tim TurnerDarrell Hughes100% (38)
- This Is A Must ReadDocument32 pagesThis Is A Must Readaeonpi97% (32)
- DA ZCR CalculatorDocument19 pagesDA ZCR CalculatorDarrell HughesNo ratings yet
- 2 Cent MailDocument13 pages2 Cent MailDarrell Hughes87% (15)
- Design Calculation of Working Platform-1Document26 pagesDesign Calculation of Working Platform-1Khandaker Khairul Alam100% (1)
- Gentry 1982 Neotropical Floristic DiversityDocument38 pagesGentry 1982 Neotropical Floristic DiversityJonathan OrtegaNo ratings yet
- Affidavit of Revocation and Rescission SSNDocument9 pagesAffidavit of Revocation and Rescission SSNDarrell Hughes100% (6)
- Overview of Functions and Operations BSPDocument2 pagesOverview of Functions and Operations BSPKarla GalvezNo ratings yet
- Bio GeographyDocument17 pagesBio Geographyindula123No ratings yet
- Bernal 2011Document10 pagesBernal 2011jazminnhalauraNo ratings yet
- Fengjiang Li, 2008Document9 pagesFengjiang Li, 2008irwanNo ratings yet
- Derryberry Etal 2011 FurnaridaeDocument14 pagesDerryberry Etal 2011 FurnaridaeNayla NascimentoNo ratings yet
- The Way the Wind Blows: Climate Change, History, and Human ActionFrom EverandThe Way the Wind Blows: Climate Change, History, and Human ActionNo ratings yet
- El Zaatari and Hublin 2014Document12 pagesEl Zaatari and Hublin 2014Aizada AlmazbekovaNo ratings yet
- Publication 4Document7 pagesPublication 4Luis Ruiz AlcántaraNo ratings yet
- Olsen Whiteside Huber 03Document42 pagesOlsen Whiteside Huber 03Juan Camilo Torres LassoNo ratings yet
- Fletcher Et Al 2013 LQ Foliar Physiogomy PDFDocument8 pagesFletcher Et Al 2013 LQ Foliar Physiogomy PDFThe PubcatNo ratings yet
- Paleoclimatic Control Archaen ProterozoicDocument21 pagesPaleoclimatic Control Archaen ProterozoicJuliano LimaNo ratings yet
- TJBDocument37 pagesTJBGabriel DonosoNo ratings yet
- Elevated Atmospheric CO and The Delayed Biotic Recovery From The End-Permian Mass ExtinctionDocument12 pagesElevated Atmospheric CO and The Delayed Biotic Recovery From The End-Permian Mass Extinctionapi-3713202No ratings yet
- Book Review of The Northern Titicaca BasDocument8 pagesBook Review of The Northern Titicaca BasThomas IossifidisNo ratings yet
- Environmental Change in The Great Plains: An Isotopic Record From Fossil HorsesDocument18 pagesEnvironmental Change in The Great Plains: An Isotopic Record From Fossil HorsesVictor Adrian Perez CrespoNo ratings yet
- GrosjeanMidHoloceneParadox2003 PDFDocument12 pagesGrosjeanMidHoloceneParadox2003 PDFDaniel Alejandro GomezNo ratings yet
- American J of Botany - 2011 - Graham - The Age and Diversification of Terrestrial New World Ecosystems Through CretaceousDocument16 pagesAmerican J of Botany - 2011 - Graham - The Age and Diversification of Terrestrial New World Ecosystems Through CretaceousAnthony PaucaNo ratings yet
- Impacts, Volcanism and Mass Extinction: Random Coincidence or Cause and Effect?Document34 pagesImpacts, Volcanism and Mass Extinction: Random Coincidence or Cause and Effect?JuanLjNo ratings yet
- Palaeogeography, Palaeoclimatology, PalaeoecologyDocument24 pagesPalaeogeography, Palaeoclimatology, Palaeoecologynlockman8No ratings yet
- Volcanically-Induced Environmental and Floral Changes Across The Triassic-Jurassic (T-J) TransitionDocument17 pagesVolcanically-Induced Environmental and Floral Changes Across The Triassic-Jurassic (T-J) TransitionDiego MagalhãesNo ratings yet
- Fig Aarons - Dust Transport To The Taylor Glacier, Antarctica, During The Last Interglacial - 2019Document10 pagesFig Aarons - Dust Transport To The Taylor Glacier, Antarctica, During The Last Interglacial - 2019mu cNo ratings yet
- Chapter - 8 Geología de ChileDocument55 pagesChapter - 8 Geología de ChilebeaparedesNo ratings yet
- Azuma Et Al 2001Document12 pagesAzuma Et Al 2001tio_conejolsvNo ratings yet
- Hannold Et Al (2021) - Mammalian Diets & Environment in Yepómera (Early Pliocene) PDFDocument16 pagesHannold Et Al (2021) - Mammalian Diets & Environment in Yepómera (Early Pliocene) PDFArturo Palma RamírezNo ratings yet
- Environmental 3Document10 pagesEnvironmental 3Thomas IossifidisNo ratings yet
- The End-Permian Extinction Glenn Simonelli Indiana UniversityDocument27 pagesThe End-Permian Extinction Glenn Simonelli Indiana Universityaneesh sebastianNo ratings yet
- Late Cretaceous Cenomanian Turonian MacrDocument69 pagesLate Cretaceous Cenomanian Turonian MacrMilan StepanovNo ratings yet
- Correlação Clima e TerraçosDocument23 pagesCorrelação Clima e TerraçosJoão Carlos CerqueiraNo ratings yet
- Restrepo Et Al - 2009 - Long-Term Erosion and Exhumation of The "Altiplano Antioqueño - EPSLDocument12 pagesRestrepo Et Al - 2009 - Long-Term Erosion and Exhumation of The "Altiplano Antioqueño - EPSLKennedy AriasNo ratings yet
- Final Draft 3Document100 pagesFinal Draft 3api-424436441No ratings yet
- Salar de Atacama BasinDocument3 pagesSalar de Atacama BasinÁlvaro Sarmiento FloresNo ratings yet
- Ecosphere - 2016 - Mizel - Rapidly Shifting Elevational Distributions of Passerine Species Parallel Vegetation Change in PDFDocument15 pagesEcosphere - 2016 - Mizel - Rapidly Shifting Elevational Distributions of Passerine Species Parallel Vegetation Change in PDFSaraNo ratings yet
- Keller 1986Document27 pagesKeller 1986dj_blade_07No ratings yet
- Schimbarile PaleoclimaticeDocument45 pagesSchimbarile PaleoclimaticeNiki VranceanuNo ratings yet
- Publicacoes Cody10 TheGreatAmericanBioticInterchangerevisitedDocument7 pagesPublicacoes Cody10 TheGreatAmericanBioticInterchangerevisitedCrocodilo VasconcelosNo ratings yet
- Unesco - Eolss Sample Chapters: Reconstructing EnvironmentsDocument6 pagesUnesco - Eolss Sample Chapters: Reconstructing EnvironmentsAngelina RandaNo ratings yet
- J Quaternary Science - 2005 - Maldonado - Pollen Analyses From A 50 000 Yr Rodent Midden Series in The Southern AtacamaDocument15 pagesJ Quaternary Science - 2005 - Maldonado - Pollen Analyses From A 50 000 Yr Rodent Midden Series in The Southern AtacamaMARÍA KORTENo ratings yet
- The Eocene - Oligocene Transition: Possible Causes, Resulting Mass Extinctions and AridificationDocument8 pagesThe Eocene - Oligocene Transition: Possible Causes, Resulting Mass Extinctions and Aridificationapi-302668669No ratings yet
- Batalha-Filho 2012 Connections Between Atlantic and Amazonian Forest Avifaunas Represent Distinct Historical EventsDocument10 pagesBatalha-Filho 2012 Connections Between Atlantic and Amazonian Forest Avifaunas Represent Distinct Historical EventsbabinechaNo ratings yet
- Cretaceous Research: G. Keller, T. Adatte, Z. Berner, E.H. Chellai, D. StuebenDocument19 pagesCretaceous Research: G. Keller, T. Adatte, Z. Berner, E.H. Chellai, D. StuebenNayanjyoti SahooNo ratings yet
- Explain The Plate Tectonics Theory.: Turcotte & Schubert, 2002)Document5 pagesExplain The Plate Tectonics Theory.: Turcotte & Schubert, 2002)Criztin Mari RochaNo ratings yet
- Vimeux & Ginot, 2006Document6 pagesVimeux & Ginot, 2006DanielNo ratings yet
- A Pulse of Mid-Pleistocene Rift Volcanism in EthioDocument12 pagesA Pulse of Mid-Pleistocene Rift Volcanism in Ethiohajali110No ratings yet
- Wastelands of Tropical Pangea: High Heat in The Permian: Neil J. TaborDocument2 pagesWastelands of Tropical Pangea: High Heat in The Permian: Neil J. TaborrikoirwantoNo ratings yet
- Ramos&Folguera09 FlatSlabSubdDocument25 pagesRamos&Folguera09 FlatSlabSubdinvarivaNo ratings yet
- Cambio Climático y EcologíaDocument12 pagesCambio Climático y EcologíaMiguel AntunezNo ratings yet
- Hillesheim, 2005Document14 pagesHillesheim, 2005Gian Pier ValenzuelaNo ratings yet
- Cretaceous Oceanography and ForaminiferaDocument29 pagesCretaceous Oceanography and ForaminiferadzzzNo ratings yet
- ChavezHoffmeister 2020 SAm FaunaDocument20 pagesChavezHoffmeister 2020 SAm FaunaLaura Sofia Chinchilla MoralesNo ratings yet
- Wen, J. Et Al. (2013) - Biogeography, Where Do We Go From HereDocument16 pagesWen, J. Et Al. (2013) - Biogeography, Where Do We Go From HereJuan David Rodriguez HurtadoNo ratings yet
- Biogeography: Where Do We Go From Here?: Jun Wen, 1 Richard H. Ree, 2 Stefanie M. Ickert-Bond, 3 Zelong Nie4 & Vicki Funk1Document16 pagesBiogeography: Where Do We Go From Here?: Jun Wen, 1 Richard H. Ree, 2 Stefanie M. Ickert-Bond, 3 Zelong Nie4 & Vicki Funk1FrontiersNo ratings yet
- Kennet T 2007Document17 pagesKennet T 2007Nayelitzin J. CanoNo ratings yet
- Wethey Et Al 2011Document13 pagesWethey Et Al 2011Princy CaesarNo ratings yet
- Clapham 2003Document18 pagesClapham 2003Hernan Gerley Alvarado RodriguezNo ratings yet
- Russoetal 2022MaltaSt PeterPoolDocument17 pagesRussoetal 2022MaltaSt PeterPoolMajed TurkistaniNo ratings yet
- Clima DEVONIANO TRIASSICODocument6 pagesClima DEVONIANO TRIASSICOWilliam MatiazziNo ratings yet
- Moffa-S-nchez Et Al-2019-Paleoceanography and PaleoclimatologyDocument53 pagesMoffa-S-nchez Et Al-2019-Paleoceanography and PaleoclimatologyDrapherNo ratings yet
- Rosenau Journal of Sedimentary Research 2013Document33 pagesRosenau Journal of Sedimentary Research 2013Andrés David VelásquezNo ratings yet
- Dust Bowl (Fuente Causas)Document25 pagesDust Bowl (Fuente Causas)LuigiNo ratings yet
- Triassic Life on Land: The Great TransitionFrom EverandTriassic Life on Land: The Great TransitionRating: 4 out of 5 stars4/5 (1)
- Invoice INV/2021/05/1122: Invoice Date: Due Date: SourceDocument1 pageInvoice INV/2021/05/1122: Invoice Date: Due Date: SourceDarrell HughesNo ratings yet
- Determining The Optimum Addition of Vegetable Materials in Adobe BricksDocument7 pagesDetermining The Optimum Addition of Vegetable Materials in Adobe BricksDarrell HughesNo ratings yet
- FM137 Mighty Mule Keypad g3Document8 pagesFM137 Mighty Mule Keypad g3Darrell HughesNo ratings yet
- Fixture Count Estimator: InstructionsDocument2 pagesFixture Count Estimator: InstructionsDarrell HughesNo ratings yet
- CDC Hearing ReportDocument39 pagesCDC Hearing ReportDarrell HughesNo ratings yet
- How To Search The SEC For A Securitized TrustDocument7 pagesHow To Search The SEC For A Securitized TrustDarrell Hughes100% (7)
- Direct Drive Smart Drive MotorsDocument9 pagesDirect Drive Smart Drive MotorsDarrell HughesNo ratings yet
- A Vertical Wind Generator From Washing Machine MotorDocument18 pagesA Vertical Wind Generator From Washing Machine MotorDarrell Hughes100% (1)
- Scope of Work Environmental Aspects Arsenic From Poultry Litter ApprovedDocument5 pagesScope of Work Environmental Aspects Arsenic From Poultry Litter ApprovedDarrell HughesNo ratings yet
- Crown of Thorns PlantDocument6 pagesCrown of Thorns PlantDarrell Hughes100% (1)
- Cancer CureDocument10 pagesCancer CureDarrell HughesNo ratings yet
- SECTION 2 Tim TurnerDocument29 pagesSECTION 2 Tim TurnerDarrell Hughes100% (15)
- HRV CaughtRedHandedDocument5 pagesHRV CaughtRedHandedDarrell Hughes100% (1)
- Pharos of HarasyDocument1 pagePharos of HarasyDarrell HughesNo ratings yet
- Basic BallisticsDocument18 pagesBasic BallisticsDarrell HughesNo ratings yet
- Ownership Structure of The Federal ReserveDocument8 pagesOwnership Structure of The Federal ReserveDarrell HughesNo ratings yet
- Instructions For Filing and Serving Demand For Bill of ParticularsDocument7 pagesInstructions For Filing and Serving Demand For Bill of ParticularsDarrell Hughes100% (2)
- Notary Acknowledgement Page in Doc FormatDocument1 pageNotary Acknowledgement Page in Doc FormatDarrell HughesNo ratings yet
- 1st Grade VocabularyDocument1 page1st Grade VocabularyamirNo ratings yet
- 3A Demand EstimationlDocument109 pages3A Demand EstimationlAlok ShuklaNo ratings yet
- ENV & MAP FileDocument43 pagesENV & MAP FileJit MukherheeNo ratings yet
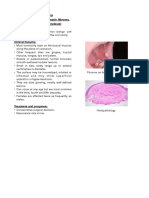
- FibromaDocument3 pagesFibromaAsiyath HNo ratings yet
- Grid ACS L2 Mobile Substations 0311 2016 03 enDocument12 pagesGrid ACS L2 Mobile Substations 0311 2016 03 enJose ValdiviesoNo ratings yet
- Term 1 Term 2 April OctoberDocument9 pagesTerm 1 Term 2 April OctoberSuleiman HaqNo ratings yet
- Compilation On Clifford Geertz TeachingsDocument25 pagesCompilation On Clifford Geertz TeachingsRobby Jane SalveronNo ratings yet
- Automotive Servicing NC I CG - Spideylab - Com - 2017Document45 pagesAutomotive Servicing NC I CG - Spideylab - Com - 2017Cristy Pearl S. GarciaNo ratings yet
- Introductory Booklet On Solar Energy For Coaching StockDocument46 pagesIntroductory Booklet On Solar Energy For Coaching StockVenkatesh VenkateshNo ratings yet
- Assessment: Facial ShavingDocument2 pagesAssessment: Facial ShavingA CNo ratings yet
- Advanced 250 V Three-Phase BLDC Controller With Embedded STM32 MCUDocument32 pagesAdvanced 250 V Three-Phase BLDC Controller With Embedded STM32 MCUOne SpringNo ratings yet
- ROLLAND ZAT540eDocument17 pagesROLLAND ZAT540egesssmNo ratings yet
- CH 02Document48 pagesCH 02miathegirl9No ratings yet
- III Eee I Sem Cs Lab Manual (Ee332) - 1Document51 pagesIII Eee I Sem Cs Lab Manual (Ee332) - 1Syed Salman AhmadNo ratings yet
- Sadhana Chatushtaya - Detailed DescriptionDocument50 pagesSadhana Chatushtaya - Detailed DescriptionSwami AjatanandaNo ratings yet
- Part 4 Crop Protection Q ADocument54 pagesPart 4 Crop Protection Q AEleven HopperNo ratings yet
- DVP-10SX PLC DeltaDocument2 pagesDVP-10SX PLC Deltawilfredomolina100% (1)
- Dust CleanDocument2 pagesDust CleanArdino Putra PerbawaNo ratings yet
- Report Project 2Document12 pagesReport Project 2Fiq IFTNo ratings yet
- Albert GerberDocument3 pagesAlbert Gerberhot_teeth100% (1)
- DIM100D283490 (1) - 7.5MVA - CommentsDocument1 pageDIM100D283490 (1) - 7.5MVA - CommentsnisarbashaNo ratings yet
- STS1 1Document42 pagesSTS1 1Peter Paul Rebucan PerudaNo ratings yet
- 11 Physics Chapter 14 and 15 Assignment 5Document2 pages11 Physics Chapter 14 and 15 Assignment 5Mohd UvaisNo ratings yet
- Tonga Tsunami Before and After Eruption - BBC NewsDocument13 pagesTonga Tsunami Before and After Eruption - BBC Newslae uneNo ratings yet
- PosterDocument1 pagePosterSaad Ullah100% (2)
- Full Metal Crown ConstructionDocument48 pagesFull Metal Crown ConstructionAnonymous k8rDEsJsU1100% (1)
- Review of Pipeline Integrity ManagementDocument8 pagesReview of Pipeline Integrity ManagementTaib Anwar100% (2)
- NPN TransistorDocument1 pageNPN TransistorsphinxNo ratings yet