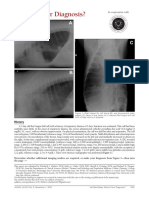
Download as pdf or txt
You might also like
- Lab Exercise 2Document4 pagesLab Exercise 2Angela Robles0% (1)
- Risk Assessment (Snake Bites, Scorpion Sting, Reptiles Bites)Document3 pagesRisk Assessment (Snake Bites, Scorpion Sting, Reptiles Bites)Raza Muhammad Soomro100% (1)
- 96-FBI Criminal Report RequestDocument2 pages96-FBI Criminal Report RequestJamesComeyJustaBitch100% (1)
- Game Snakes Ladders With English Game Questions and InstructionsDocument10 pagesGame Snakes Ladders With English Game Questions and InstructionsquadromagicoNo ratings yet
- AnimalClassificationandAnimalCharacteristicsSortsFREEBIE PDFDocument8 pagesAnimalClassificationandAnimalCharacteristicsSortsFREEBIE PDFtanleeleanNo ratings yet
- Historical Perspective: A Decade of Discoveries in Cardiac BiologyDocument8 pagesHistorical Perspective: A Decade of Discoveries in Cardiac BiologyDiego AbrahamNo ratings yet
- Developmental Biology: The Epicardium in The Embryonic and Adult ZebrafishDocument16 pagesDevelopmental Biology: The Epicardium in The Embryonic and Adult ZebrafishplatusNo ratings yet
- Embryology of The Hind GutDocument10 pagesEmbryology of The Hind Gutproject-247758No ratings yet
- Nature 2014 Long JADocument13 pagesNature 2014 Long JAAlfredo PerettiNo ratings yet
- Cardiogenesis and The Regulation of Cardiac-Specific Gene ExpressionDocument14 pagesCardiogenesis and The Regulation of Cardiac-Specific Gene ExpressiontyasNo ratings yet
- Biodesarrollo Corazon ReptilesDocument12 pagesBiodesarrollo Corazon ReptilesAlex Ramos maqueraNo ratings yet
- 2011 BFnature09655 MOESM201 ESMDocument11 pages2011 BFnature09655 MOESM201 ESMliqinggreenNo ratings yet
- Identification Guide To Antarctic CopepodsDocument42 pagesIdentification Guide To Antarctic CopepodsmauricioNo ratings yet
- Metabolic Control of Progenitor Cell Propagation During Drosophila Tracheal RemodelingDocument17 pagesMetabolic Control of Progenitor Cell Propagation During Drosophila Tracheal Remodelingdede chenNo ratings yet
- Journal of Morphology - 2019 - Kroneman - Comparative Analysis of Avian Hearts Provides Little Evidence For Variation AmongDocument16 pagesJournal of Morphology - 2019 - Kroneman - Comparative Analysis of Avian Hearts Provides Little Evidence For Variation AmongJohanne May Capitulo LopezNo ratings yet
- Ectopic Thyroid: Etiology, Pathology and Management: ReviewDocument9 pagesEctopic Thyroid: Etiology, Pathology and Management: ReviewIonuț RusNo ratings yet
- tmp75B6 TMPDocument4 pagestmp75B6 TMPFrontiersNo ratings yet
- Structure and Function of The Developing Zebrafish HeartDocument10 pagesStructure and Function of The Developing Zebrafish HeartKarthik ShettyNo ratings yet
- tmp3FED TMPDocument3 pagestmp3FED TMPFrontiersNo ratings yet
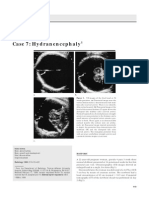
- Case 7: Hydranencephaly: Alfred B. Kurtz, MD Pamela T. Johnson, MDDocument4 pagesCase 7: Hydranencephaly: Alfred B. Kurtz, MD Pamela T. Johnson, MDKenzo Adhi WiranataNo ratings yet
- Lobulated Spleen With A Fissure On Diaphragmatic Surface: RD RDDocument7 pagesLobulated Spleen With A Fissure On Diaphragmatic Surface: RD RDShantu ShirurmathNo ratings yet
- Frog EmbryosDocument43 pagesFrog EmbryosNics Martinez100% (7)
- 560 FTPDocument8 pages560 FTPSrinivas PrasadNo ratings yet
- Kleckowska Et Al Macroanatomical Investigation of ARGDocument5 pagesKleckowska Et Al Macroanatomical Investigation of ARGGNo ratings yet
- Embryology of Oesophageal Atresia: Adonis S. Ioannides, Andrew J. CoppDocument10 pagesEmbryology of Oesophageal Atresia: Adonis S. Ioannides, Andrew J. CoppXimenita GhilardiNo ratings yet
- Structure of The Reproductive Apparatus and Life Cycle of Milax Gagates Draparnaud Mollusca Gastropoda PulmonataDocument40 pagesStructure of The Reproductive Apparatus and Life Cycle of Milax Gagates Draparnaud Mollusca Gastropoda PulmonataRaihana Naifa ENo ratings yet
- Tetralogy of Fallot, Agarwala 2017Document5 pagesTetralogy of Fallot, Agarwala 2017rinayondaNo ratings yet
- Maduracion de La AmigadalaDocument15 pagesMaduracion de La AmigadalaAracelis Calzadilla NúñezNo ratings yet
- E7.5 E8.0 E8.5 E12.5Document11 pagesE7.5 E8.0 E8.5 E12.5خـޢޢـطـޢޢـ آحـޢޢـمـޢޢـرNo ratings yet
- Ancient Animal MicroRNAs and The Evolution of Tissue IdentityDocument10 pagesAncient Animal MicroRNAs and The Evolution of Tissue IdentityRebecca PotterNo ratings yet
- Comisuritis Posterior 2Document8 pagesComisuritis Posterior 2VORTIZNo ratings yet
- 2 PDFDocument8 pages2 PDFanon_243369186No ratings yet
- AVIAN-The Avıan Spleen Anatomy, Physiology and DiagnosticsDocument6 pagesAVIAN-The Avıan Spleen Anatomy, Physiology and Diagnosticstaner_soysuren100% (1)
- Developmental Dynamics - 2006 - Campione - Cardiovascular Development Toward Biomedical ApplicabilityDocument3 pagesDevelopmental Dynamics - 2006 - Campione - Cardiovascular Development Toward Biomedical Applicabilitystevenburrow06No ratings yet
- Congenital LaryngomalaciaDocument8 pagesCongenital LaryngomalaciaRettha SigiroNo ratings yet
- Tiroida Ectopica 1Document8 pagesTiroida Ectopica 1Ionuț RusNo ratings yet
- Sall1 Maintains Nephron Progenitors and NascentDocument12 pagesSall1 Maintains Nephron Progenitors and NascentWilan KrisnaNo ratings yet
- 2012cooper Et Al Wires Development of Bat FlightDocument7 pages2012cooper Et Al Wires Development of Bat Flightapi-237346249No ratings yet
- Principles of DevelopmentDocument9 pagesPrinciples of Developmentsoccerrocksj18No ratings yet
- JHG 201710Document5 pagesJHG 201710maow2No ratings yet
- Hendler 1986Document8 pagesHendler 1986byhz7wwdm4No ratings yet
- Journal of Morphology SpermwarteryDocument14 pagesJournal of Morphology SpermwarteryJustin RamonNo ratings yet
- Baf60mice Takeuchi2007Document7 pagesBaf60mice Takeuchi2007vweakeNo ratings yet
- Zool Lab 4Document17 pagesZool Lab 4pascualfrancejosephNo ratings yet
- Establishment of Vertebrate Left-Right Asymmetry: Hiroshi Hamada, Chikara Meno, Daisuke Watanabe and Yukio SaijohDocument11 pagesEstablishment of Vertebrate Left-Right Asymmetry: Hiroshi Hamada, Chikara Meno, Daisuke Watanabe and Yukio SaijohDianne AcobaNo ratings yet
- Anani Et Al 2014Document12 pagesAnani Et Al 2014nugrahoneyNo ratings yet
- Vanderputte 2011Document8 pagesVanderputte 2011adamNo ratings yet
- Anomalous Development of Thyroid Gland A Cadaveric Study in Coastal Population of Andhra PradeshDocument4 pagesAnomalous Development of Thyroid Gland A Cadaveric Study in Coastal Population of Andhra Pradeshتقوى اللهNo ratings yet
- Description of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Document13 pagesDescription of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Kanhiya MahourNo ratings yet
- Tetralogía en Hurón - CASODocument6 pagesTetralogía en Hurón - CASOLAURA AILED LOPEZ LOZANo ratings yet
- The Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneDocument13 pagesThe Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneCandle Jo'inNo ratings yet
- Developmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellDocument3 pagesDevelopmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellAroa ChansNo ratings yet
- Neuro 2Document5 pagesNeuro 2Ivan RisnaNo ratings yet
- TMP AABEDocument2 pagesTMP AABEFrontiersNo ratings yet
- Garces 2007Document6 pagesGarces 2007Peter CordovageNo ratings yet
- Pi Is 1878540914000462Document6 pagesPi Is 1878540914000462aandakuNo ratings yet
- Articulo de EmbrioDocument10 pagesArticulo de EmbrioGabriel Pineda CastroNo ratings yet
- Neuroanatomy of The Zebrafish Brain - A Topological Atlas 1996Document141 pagesNeuroanatomy of The Zebrafish Brain - A Topological Atlas 1996Daniel RuizNo ratings yet
- Splina Functii PDFDocument11 pagesSplina Functii PDFTotolici StefanNo ratings yet
- Stenotic Semilunar Valve in Persistent Truncus ArteriosusDocument6 pagesStenotic Semilunar Valve in Persistent Truncus ArteriosusAlma Grace Pineda TeránNo ratings yet
- Akash Aipmt Final 2011 SolutionDocument18 pagesAkash Aipmt Final 2011 Solutionsaurav guptaNo ratings yet
- Genes Regulating Lymphangiogenesis Control Venous Valve Formation and Maintenance in MiceDocument10 pagesGenes Regulating Lymphangiogenesis Control Venous Valve Formation and Maintenance in MiceGerardo OrtizNo ratings yet
- Ajpendo 00705 2006Document8 pagesAjpendo 00705 2006Fujiko SaavedraNo ratings yet
- 93 Welcome To The Revolution AgendaDocument1 page93 Welcome To The Revolution AgendaJamesComeyJustaBitch0% (2)
- 87-Triune Brain TheoryDocument7 pages87-Triune Brain TheoryJamesComeyJustaBitchNo ratings yet
- 86-Tp Scales TalesDocument19 pages86-Tp Scales TalesJamesComeyJustaBitchNo ratings yet
- The Evolution of The Amygdala: Basic FunctionDocument8 pagesThe Evolution of The Amygdala: Basic FunctionJamesComeyJustaBitchNo ratings yet
- 83-The Triune BrainDocument4 pages83-The Triune BrainJamesComeyJustaBitchNo ratings yet
- Suffixes: Introduction: Suffixes Are Syllables Added To The Ends of Words To Change Their Functions. For ExampleDocument5 pagesSuffixes: Introduction: Suffixes Are Syllables Added To The Ends of Words To Change Their Functions. For ExampleJamesComeyJustaBitchNo ratings yet
- Reptilian All The Way?: LetterDocument1 pageReptilian All The Way?: LetterJamesComeyJustaBitchNo ratings yet
- Avian and Reptilian Evaporative CompartmentalizationDocument6 pagesAvian and Reptilian Evaporative CompartmentalizationJamesComeyJustaBitchNo ratings yet
- 73-Reptilian Abductions in The..Document3 pages73-Reptilian Abductions in The..JamesComeyJustaBitchNo ratings yet
- 67 ParasiteIdentificationDocument1 page67 ParasiteIdentificationJamesComeyJustaBitchNo ratings yet
- 78 Reptillian AggDocument1 page78 Reptillian AggJamesComeyJustaBitchNo ratings yet
- 74-Reptilian Agenda Milky Way Galaxy Catastrophe Year 2370Document5 pages74-Reptilian Agenda Milky Way Galaxy Catastrophe Year 2370JamesComeyJustaBitch100% (1)
- 60 Marshall ArticleDocument7 pages60 Marshall ArticleJamesComeyJustaBitchNo ratings yet
- Environmental Persistence of Amphibian and Reptilian RanavirusesDocument8 pagesEnvironmental Persistence of Amphibian and Reptilian RanavirusesJamesComeyJustaBitchNo ratings yet
- 59 MARS Andrew D Basiago Reptoid Predation Found On Mars 4-12-09Document5 pages59 MARS Andrew D Basiago Reptoid Predation Found On Mars 4-12-09JamesComeyJustaBitchNo ratings yet
- Human Language and Our Reptilian Brain: The Subcortical Bases of Speech, Syntax, and ThoughtDocument20 pagesHuman Language and Our Reptilian Brain: The Subcortical Bases of Speech, Syntax, and ThoughtJamesComeyJustaBitchNo ratings yet
- 031 Bahasa - Innggris - 1Document28 pages031 Bahasa - Innggris - 1Tavip ZulaifahNo ratings yet
- Evolution of Respiratory SystemDocument10 pagesEvolution of Respiratory SystemWwwanand111No ratings yet
- The Animal Kingdom: Living On Planet EarthDocument19 pagesThe Animal Kingdom: Living On Planet EarthSri Latha Ganesh RaoNo ratings yet
- Evolution, Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution, Epidemiology and Etiology of Temporomandibular Joint DisordersakNo ratings yet
- E.V.O Search For Eden DetonadoDocument51 pagesE.V.O Search For Eden DetonadoAlan KardecNo ratings yet
- Lola The Animal Helper ScienceDocument27 pagesLola The Animal Helper ScienceCryptoS hitNo ratings yet
- Fossils in Utah Booklet 1Document21 pagesFossils in Utah Booklet 1api-218354277No ratings yet
- Comparative Placentation: GlossaryDocument8 pagesComparative Placentation: GlossaryJosh Soriano IiNo ratings yet
- One Million Things Animal LifeDocument127 pagesOne Million Things Animal Lifesebibv100% (5)
- 2nd Quarter Summative Test No.2Document2 pages2nd Quarter Summative Test No.2Marilou KimayongNo ratings yet
- Declarative Programming ExercisesDocument19 pagesDeclarative Programming ExercisesadaNo ratings yet
- Unit 2 Lesson 8 Super Sea Animals CLIL ScienceDocument29 pagesUnit 2 Lesson 8 Super Sea Animals CLIL Sciencemylyn ortizNo ratings yet
- BSC Zoology Part II ReptiliaDocument5 pagesBSC Zoology Part II Reptiliayasahswi91No ratings yet
- 8-Bit Adventures - Space Bounty Hunters PDFDocument48 pages8-Bit Adventures - Space Bounty Hunters PDFJohn Churchill100% (1)
- Life-History Evolution in ReptilesDocument27 pagesLife-History Evolution in ReptilesPahal AgarwalNo ratings yet
- Classification of AnimalsDocument33 pagesClassification of Animalsrajeshmmahajan100% (1)
- PT - SCIENCE 6 - Q2 - FinalDocument5 pagesPT - SCIENCE 6 - Q2 - FinalIrine BrionesNo ratings yet
- An Updated Checklist of The Amphibians and Reptiles of NicaraguaDocument18 pagesAn Updated Checklist of The Amphibians and Reptiles of NicaraguaRicardo SozaNo ratings yet
- O P Jindal Modern School, Hisar: Worksheet No. 3 Subject-Evs NAME: ................... ROLL NO. . Class 5Document2 pagesO P Jindal Modern School, Hisar: Worksheet No. 3 Subject-Evs NAME: ................... ROLL NO. . Class 5Raghav BansalNo ratings yet
- DETAILED LESSON PLAN Hierachical System ClassificationsDocument22 pagesDETAILED LESSON PLAN Hierachical System ClassificationsMary Rose RamosNo ratings yet
- Natural HistoryDocument664 pagesNatural Historykiradiologija75% (4)
- PTS 2019 2020 40 Nomer SoalDocument6 pagesPTS 2019 2020 40 Nomer SoalRizki HariyantiNo ratings yet
- Unit 2 Exploring The Living World: Multiple-Choice QuestionsDocument18 pagesUnit 2 Exploring The Living World: Multiple-Choice QuestionsVincent NGNo ratings yet
- REPTILESDocument2 pagesREPTILESCharll Dezon Abcede AbaNo ratings yet
- A Detailed Lesson Plan in Mathematic VI g4 and 6Document12 pagesA Detailed Lesson Plan in Mathematic VI g4 and 6Ivy Novero SibuloNo ratings yet
- Tuatara-Climate-Change StudentbDocument5 pagesTuatara-Climate-Change Studentbapi-406290658No ratings yet
- Tugas Bahasa InggrisDocument5 pagesTugas Bahasa InggrisVvyynxNo ratings yet