
Emb Oj 2012146 A
Emb Oj 2012146 A
You might also like
- Hhmi Cell Cycle and Cancer - CompletedDocument3 pagesHhmi Cell Cycle and Cancer - CompletedAbel89% (9)
- Test Bank For Essentials of Biology 5th Edition by MaderDocument36 pagesTest Bank For Essentials of Biology 5th Edition by Madermichellesmallmatfqcendg100% (28)
- Paul Schmid-Hempel - Evolutionary Parasitology - The Integrated Study of Infections, Immunology, Ecology, and Genetics-Oxford University Press (2021)Document572 pagesPaul Schmid-Hempel - Evolutionary Parasitology - The Integrated Study of Infections, Immunology, Ecology, and Genetics-Oxford University Press (2021)Guilherme JungNo ratings yet
- Gbbe Chapter 2 CancerDocument20 pagesGbbe Chapter 2 Cancerapi-374854456No ratings yet
- History of CancerDocument17 pagesHistory of CancercheaussieNo ratings yet
- Patho Ch. 10-12Document13 pagesPatho Ch. 10-12JoyyNo ratings yet
- Is Virology Dead?Document3 pagesIs Virology Dead?gabrielgpinto1No ratings yet
- BMC Evolutionary Biology: Giant Viruses, Giant Chimeras: The Multiple Evolutionary Histories of Mimivirus GenesDocument10 pagesBMC Evolutionary Biology: Giant Viruses, Giant Chimeras: The Multiple Evolutionary Histories of Mimivirus GenesKaio FernandesNo ratings yet
- Advances in Parasitology, Vol. 48 (B-Ok - Xyz)Document407 pagesAdvances in Parasitology, Vol. 48 (B-Ok - Xyz)Jose Franco NaviaNo ratings yet
- Papillomaviruses - Viral Evolution, Cancer and Evolutionary MedicineDocument20 pagesPapillomaviruses - Viral Evolution, Cancer and Evolutionary MedicineWilliam PeiNo ratings yet
- The Origins of VirusesDocument4 pagesThe Origins of VirusesStiven BFNo ratings yet
- b76eeVIRUSES 4Document4 pagesb76eeVIRUSES 4Mudit MisraNo ratings yet
- Dissertation Sur Le Virus EbolaDocument5 pagesDissertation Sur Le Virus EbolaCustomPaperServiceCanada100% (1)
- The Origins of VirusesDocument6 pagesThe Origins of VirusesKevin van HollandNo ratings yet
- Coming Full Circle-From Endless Complexity To Simplicity and Back AgainDocument5 pagesComing Full Circle-From Endless Complexity To Simplicity and Back AgainMariano PerezNo ratings yet
- 2024-Nature Ecology & EvolutionDocument2 pages2024-Nature Ecology & EvolutionIgnacio martinNo ratings yet
- Virology: James M. PipasDocument10 pagesVirology: James M. PipasOday AdaNo ratings yet
- History of Arbovirology - Memories From The Field Volume II - Virus Family and Regional Perspectives, Molecular Biology and PathogenesisDocument535 pagesHistory of Arbovirology - Memories From The Field Volume II - Virus Family and Regional Perspectives, Molecular Biology and PathogenesisItrux jonathan cisnerosNo ratings yet
- Evolutionary Parasitology The Integrated Study of Infections Immunology Ecology and Genetics 2Nd Edition Paul Schmid Hempel Full ChapterDocument67 pagesEvolutionary Parasitology The Integrated Study of Infections Immunology Ecology and Genetics 2Nd Edition Paul Schmid Hempel Full Chaptervivian.arriola772100% (4)
- p53 My WorkbookDocument12 pagesp53 My WorkbookArunanshu PalNo ratings yet
- Nature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetenceDocument30 pagesNature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetencedianamfgNo ratings yet
- Evolution of VirusesDocument5 pagesEvolution of VirusesYolo WoloNo ratings yet
- Viruses 12 01341Document10 pagesViruses 12 01341gabrielgpinto1No ratings yet
- Footprints of Past Pandemics in The Human Genome - OcredDocument7 pagesFootprints of Past Pandemics in The Human Genome - OcredAndersinclair AndersinclairNo ratings yet
- Chinhoyi University of Technology: Biotechnology DepartmentDocument8 pagesChinhoyi University of Technology: Biotechnology DepartmentDumisani Nguni100% (1)
- Thannesberger 2017Document15 pagesThannesberger 2017Денис КрахоткинNo ratings yet
- Bottlenose Dolphin (Tursiops Truncatus) Papillomaviruses: Vaccine Antigen Candidates and Screening Test DevelopmentDocument11 pagesBottlenose Dolphin (Tursiops Truncatus) Papillomaviruses: Vaccine Antigen Candidates and Screening Test Developmentalan_kadirNo ratings yet
- Hallmark of CancerDocument14 pagesHallmark of CanceriinsabatiniNo ratings yet
- Virology - Chapter Six Oncogenic Viruses: Classes of Tumor VirusesDocument17 pagesVirology - Chapter Six Oncogenic Viruses: Classes of Tumor VirusesMd Daud AliNo ratings yet
- The Hallmarks of Cancer Review: 2000 by Cell PressDocument14 pagesThe Hallmarks of Cancer Review: 2000 by Cell PressAashiq HussainNo ratings yet
- XMRV Slides Jan2010Document28 pagesXMRV Slides Jan2010elipoarchNo ratings yet
- Are Viruses Alive?: by Luis P. VillarrealDocument8 pagesAre Viruses Alive?: by Luis P. VillarrealTrixie SantosNo ratings yet
- Engineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyDocument6 pagesEngineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyZsi GaNo ratings yet
- Examen Final Univ Litoral Bioq-finalesIIDocument5 pagesExamen Final Univ Litoral Bioq-finalesIIKorey BoyerNo ratings yet
- HIV - Structure Life Cycle and PathogenecityDocument26 pagesHIV - Structure Life Cycle and PathogenecityhunterbomkuachNo ratings yet
- Vasilakis Et Al., 2019Document38 pagesVasilakis Et Al., 2019Lúcia ReisNo ratings yet
- هرووبیماری جدید تکرارDocument16 pagesهرووبیماری جدید تکرارkaveh.azizi24No ratings yet
- The Genetic Basis of CancerDocument8 pagesThe Genetic Basis of CancerLim ZYNo ratings yet
- Periodontal Microbial Dark MatterDocument16 pagesPeriodontal Microbial Dark MatterAndrés Nicolás Beltrán SeguraNo ratings yet
- HIV A Virus Like No OtherDocument83 pagesHIV A Virus Like No Otherbadboy6.868510No ratings yet
- Dunn RefDocument47 pagesDunn RefFlorencia Chandrika HalimNo ratings yet
- (Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springer International Publishing - Humana Press (2019)Document406 pages(Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springer International Publishing - Humana Press (2019)karima majdouliNo ratings yet
- (Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springe PDFDocument406 pages(Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springe PDFmariomgNo ratings yet
- What Is CancerDocument2 pagesWhat Is CancerDebapriya HazraNo ratings yet
- Human Pathology (2008) 39, 154-166Document13 pagesHuman Pathology (2008) 39, 154-166Laura valentina ramos mezaNo ratings yet
- First Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDocument11 pagesFirst Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDavid A. Osorio G.No ratings yet
- Origin and Cellular Evolution-Claudiu BandeaDocument16 pagesOrigin and Cellular Evolution-Claudiu BandeaMedicina Usmp Filial NorteNo ratings yet
- Viruses & EvolutionDocument27 pagesViruses & Evolutionhaluk bilginNo ratings yet
- Exosomes - The Protagonists in The Tale of Colorectal CancerDocument38 pagesExosomes - The Protagonists in The Tale of Colorectal CancerManoj Kumar pandreNo ratings yet
- Research Paper On SmallpoxDocument11 pagesResearch Paper On Smallpoxxsykcbikf100% (1)
- The Role of Human Papillomavirus in Pre-Cancerous Lesions and Oral CancersDocument29 pagesThe Role of Human Papillomavirus in Pre-Cancerous Lesions and Oral CancerssevattapillaiNo ratings yet
- Notes 1Document5 pagesNotes 1logonautNo ratings yet
- Chronic CancerDocument13 pagesChronic Cancerjulius_henryNo ratings yet
- Tropicalmed 08 00173 v2Document12 pagesTropicalmed 08 00173 v2cesar martin seguraNo ratings yet
- David S. Lindsay, Louis M. Weiss - Opportunistic Infections - Toxoplasma, Sarcocystis, and Microsporidia (World Class Parasites) (2004)Document249 pagesDavid S. Lindsay, Louis M. Weiss - Opportunistic Infections - Toxoplasma, Sarcocystis, and Microsporidia (World Class Parasites) (2004)DuONo ratings yet
- The Dna Provirus HypothesisDocument10 pagesThe Dna Provirus HypothesismarsbleachNo ratings yet
- Jackie Virology AssignmentDocument6 pagesJackie Virology AssignmentDumisani NguniNo ratings yet
- Infectious Disease Ecology of Wild Birds Jennifer C Owen Full ChapterDocument48 pagesInfectious Disease Ecology of Wild Birds Jennifer C Owen Full Chapterray.marks344100% (12)
- 10201148Document96 pages10201148Huzaifa SyedNo ratings yet
- Transformation and Oncogenesis: Biology W3310/4310 Virology Spring 2016Document56 pagesTransformation and Oncogenesis: Biology W3310/4310 Virology Spring 2016kashif manzoorNo ratings yet
- The Basics of VirusesDocument4 pagesThe Basics of VirusesSharolt SierraNo ratings yet
- From Mendel To The Human Genome Project: The Implications For Nurse EducationDocument6 pagesFrom Mendel To The Human Genome Project: The Implications For Nurse EducationKome ČemuNo ratings yet
- TOTUL Despre HPVDocument220 pagesTOTUL Despre HPVsweetchildofmineNo ratings yet
- Waqar AhmedDocument11 pagesWaqar AhmedSyed Hassan GillaniNo ratings yet
- Viruses: A Scientific American Library BookFrom EverandViruses: A Scientific American Library BookRating: 4.5 out of 5 stars4.5/5 (11)
- BT 2019Document13 pagesBT 2019biotech_vidhyaNo ratings yet
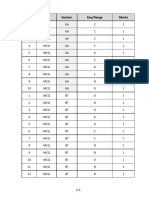
- Q.No. Type Section Key/Range MarksDocument3 pagesQ.No. Type Section Key/Range Marksbiotech_vidhyaNo ratings yet
- Troubleshooting SDS-PAGE 1Document3 pagesTroubleshooting SDS-PAGE 1biotech_vidhyaNo ratings yet
- Buffer Preparation PDFDocument6 pagesBuffer Preparation PDFbiotech_vidhyaNo ratings yet
- Whole Cell ExtractDocument1 pageWhole Cell Extractbiotech_vidhyaNo ratings yet
- Polymerase Chain Reaction (PCR)Document3 pagesPolymerase Chain Reaction (PCR)biotech_vidhyaNo ratings yet
- Nuclear ExtractsDocument2 pagesNuclear Extractsbiotech_vidhyaNo ratings yet
- Buffer Preparation PDFDocument6 pagesBuffer Preparation PDFbiotech_vidhyaNo ratings yet
- Ies 17 Set A Me Q ADocument67 pagesIes 17 Set A Me Q Abiotech_vidhyaNo ratings yet
- Mechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010Document20 pagesMechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010biotech_vidhyaNo ratings yet
- Befcv List PDFDocument22 pagesBefcv List PDFbiotech_vidhyaNo ratings yet
- TNPSC Group 1 Prelim Book List PDFDocument2 pagesTNPSC Group 1 Prelim Book List PDFbiotech_vidhyaNo ratings yet
- 1 TolerancesDocument1 page1 Tolerancesbiotech_vidhyaNo ratings yet
- Teachers Recruitment Board: 1. Important DatesDocument13 pagesTeachers Recruitment Board: 1. Important Datesbiotech_vidhyaNo ratings yet
- Isa BusDocument30 pagesIsa Busbiotech_vidhya100% (1)
- Lec09 Three PhaseDocument84 pagesLec09 Three Phasebiotech_vidhyaNo ratings yet
- K - 1 Perspective and The Future of OncologyDocument36 pagesK - 1 Perspective and The Future of OncologyPRAGAATHYRAJANNo ratings yet
- Imunologi Tumor: Syarifuddin WahidDocument40 pagesImunologi Tumor: Syarifuddin WahidinhaNo ratings yet
- Decoding Cancer - Lesson 1 - Interactive Lesson - July - 2018Document46 pagesDecoding Cancer - Lesson 1 - Interactive Lesson - July - 2018NisrinNo ratings yet
- Ch4 Lecture SlidesDocument34 pagesCh4 Lecture SlidesJad AwadNo ratings yet
- This Study Resource Was: G R A D E S L A B - C O MDocument8 pagesThis Study Resource Was: G R A D E S L A B - C O MHugsNo ratings yet
- 02 STR 9 102Document94 pages02 STR 9 10211111No ratings yet
- Neoplasia PDFDocument182 pagesNeoplasia PDFmarwaNo ratings yet
- Cancer Genetics: Sreekutty S 2 MSC ZoologyDocument34 pagesCancer Genetics: Sreekutty S 2 MSC ZoologyShamsudheen maharajasNo ratings yet
- Genomes and Their EvolutionDocument23 pagesGenomes and Their Evolutionprehealthhelp100% (2)
- Emperor of All Maladies CastDocument6 pagesEmperor of All Maladies Castmykwin07No ratings yet
- FMS 1 - Week 7 Tutorial 1 LO PDFDocument17 pagesFMS 1 - Week 7 Tutorial 1 LO PDFAprillia AlmaasNo ratings yet
- Plants That Fight CancerDocument314 pagesPlants That Fight CancerMai Elnaggar100% (4)
- Chapter 14 Mutation and CancerDocument43 pagesChapter 14 Mutation and CancerCharlieHuangNo ratings yet
- Neoplasia: Nomenclature Epidemiology OncogenesDocument7 pagesNeoplasia: Nomenclature Epidemiology OncogenespriyaNo ratings yet
- Oncogene EtcDocument48 pagesOncogene EtcInsyirah JohariNo ratings yet
- General Biology 1: Quarter 1 - Module - : Title: Cell CycleDocument27 pagesGeneral Biology 1: Quarter 1 - Module - : Title: Cell CycleRea A. Bilan0% (1)
- CarcinogenesisDocument3 pagesCarcinogenesiskishorkumarn8212No ratings yet
- Cytogenetics Final ExamDocument20 pagesCytogenetics Final ExamMICHAEL JOHN AGUILARNo ratings yet
- 6 Hos CarcinogenesisDocument90 pages6 Hos CarcinogenesisalgutNo ratings yet
- NikitaDocument41 pagesNikitaAtika AnggrainiNo ratings yet
- 1.biokimia KankerDocument94 pages1.biokimia Kankersebastian chendraNo ratings yet
- Onco, TSG & CancerDocument8 pagesOnco, TSG & Cancersumera120488No ratings yet
- Chemotherapy of Cancer: Submitted by Syama - J.S Roll No:15 2 SEM M Pharm PharmacologyDocument64 pagesChemotherapy of Cancer: Submitted by Syama - J.S Roll No:15 2 SEM M Pharm PharmacologySyama J.S100% (1)
- Quiz BankDocument12 pagesQuiz Banknadim1241335No ratings yet
- Tumor VirusesDocument28 pagesTumor VirusespavindersinghNo ratings yet








































































































Download as pdf or txt
You might also like
- Hhmi Cell Cycle and Cancer - CompletedDocument3 pagesHhmi Cell Cycle and Cancer - CompletedAbel89% (9)
- Test Bank For Essentials of Biology 5th Edition by MaderDocument36 pagesTest Bank For Essentials of Biology 5th Edition by Madermichellesmallmatfqcendg100% (28)
- Paul Schmid-Hempel - Evolutionary Parasitology - The Integrated Study of Infections, Immunology, Ecology, and Genetics-Oxford University Press (2021)Document572 pagesPaul Schmid-Hempel - Evolutionary Parasitology - The Integrated Study of Infections, Immunology, Ecology, and Genetics-Oxford University Press (2021)Guilherme JungNo ratings yet
- Gbbe Chapter 2 CancerDocument20 pagesGbbe Chapter 2 Cancerapi-374854456No ratings yet
- History of CancerDocument17 pagesHistory of CancercheaussieNo ratings yet
- Patho Ch. 10-12Document13 pagesPatho Ch. 10-12JoyyNo ratings yet
- Is Virology Dead?Document3 pagesIs Virology Dead?gabrielgpinto1No ratings yet
- BMC Evolutionary Biology: Giant Viruses, Giant Chimeras: The Multiple Evolutionary Histories of Mimivirus GenesDocument10 pagesBMC Evolutionary Biology: Giant Viruses, Giant Chimeras: The Multiple Evolutionary Histories of Mimivirus GenesKaio FernandesNo ratings yet
- Advances in Parasitology, Vol. 48 (B-Ok - Xyz)Document407 pagesAdvances in Parasitology, Vol. 48 (B-Ok - Xyz)Jose Franco NaviaNo ratings yet
- Papillomaviruses - Viral Evolution, Cancer and Evolutionary MedicineDocument20 pagesPapillomaviruses - Viral Evolution, Cancer and Evolutionary MedicineWilliam PeiNo ratings yet
- The Origins of VirusesDocument4 pagesThe Origins of VirusesStiven BFNo ratings yet
- b76eeVIRUSES 4Document4 pagesb76eeVIRUSES 4Mudit MisraNo ratings yet
- Dissertation Sur Le Virus EbolaDocument5 pagesDissertation Sur Le Virus EbolaCustomPaperServiceCanada100% (1)
- The Origins of VirusesDocument6 pagesThe Origins of VirusesKevin van HollandNo ratings yet
- Coming Full Circle-From Endless Complexity To Simplicity and Back AgainDocument5 pagesComing Full Circle-From Endless Complexity To Simplicity and Back AgainMariano PerezNo ratings yet
- 2024-Nature Ecology & EvolutionDocument2 pages2024-Nature Ecology & EvolutionIgnacio martinNo ratings yet
- Virology: James M. PipasDocument10 pagesVirology: James M. PipasOday AdaNo ratings yet
- History of Arbovirology - Memories From The Field Volume II - Virus Family and Regional Perspectives, Molecular Biology and PathogenesisDocument535 pagesHistory of Arbovirology - Memories From The Field Volume II - Virus Family and Regional Perspectives, Molecular Biology and PathogenesisItrux jonathan cisnerosNo ratings yet
- Evolutionary Parasitology The Integrated Study of Infections Immunology Ecology and Genetics 2Nd Edition Paul Schmid Hempel Full ChapterDocument67 pagesEvolutionary Parasitology The Integrated Study of Infections Immunology Ecology and Genetics 2Nd Edition Paul Schmid Hempel Full Chaptervivian.arriola772100% (4)
- p53 My WorkbookDocument12 pagesp53 My WorkbookArunanshu PalNo ratings yet
- Nature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetenceDocument30 pagesNature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission CompetencedianamfgNo ratings yet
- Evolution of VirusesDocument5 pagesEvolution of VirusesYolo WoloNo ratings yet
- Viruses 12 01341Document10 pagesViruses 12 01341gabrielgpinto1No ratings yet
- Footprints of Past Pandemics in The Human Genome - OcredDocument7 pagesFootprints of Past Pandemics in The Human Genome - OcredAndersinclair AndersinclairNo ratings yet
- Chinhoyi University of Technology: Biotechnology DepartmentDocument8 pagesChinhoyi University of Technology: Biotechnology DepartmentDumisani Nguni100% (1)
- Thannesberger 2017Document15 pagesThannesberger 2017Денис КрахоткинNo ratings yet
- Bottlenose Dolphin (Tursiops Truncatus) Papillomaviruses: Vaccine Antigen Candidates and Screening Test DevelopmentDocument11 pagesBottlenose Dolphin (Tursiops Truncatus) Papillomaviruses: Vaccine Antigen Candidates and Screening Test Developmentalan_kadirNo ratings yet
- Hallmark of CancerDocument14 pagesHallmark of CanceriinsabatiniNo ratings yet
- Virology - Chapter Six Oncogenic Viruses: Classes of Tumor VirusesDocument17 pagesVirology - Chapter Six Oncogenic Viruses: Classes of Tumor VirusesMd Daud AliNo ratings yet
- The Hallmarks of Cancer Review: 2000 by Cell PressDocument14 pagesThe Hallmarks of Cancer Review: 2000 by Cell PressAashiq HussainNo ratings yet
- XMRV Slides Jan2010Document28 pagesXMRV Slides Jan2010elipoarchNo ratings yet
- Are Viruses Alive?: by Luis P. VillarrealDocument8 pagesAre Viruses Alive?: by Luis P. VillarrealTrixie SantosNo ratings yet
- Engineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyDocument6 pagesEngineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyZsi GaNo ratings yet
- Examen Final Univ Litoral Bioq-finalesIIDocument5 pagesExamen Final Univ Litoral Bioq-finalesIIKorey BoyerNo ratings yet
- HIV - Structure Life Cycle and PathogenecityDocument26 pagesHIV - Structure Life Cycle and PathogenecityhunterbomkuachNo ratings yet
- Vasilakis Et Al., 2019Document38 pagesVasilakis Et Al., 2019Lúcia ReisNo ratings yet
- هرووبیماری جدید تکرارDocument16 pagesهرووبیماری جدید تکرارkaveh.azizi24No ratings yet
- The Genetic Basis of CancerDocument8 pagesThe Genetic Basis of CancerLim ZYNo ratings yet
- Periodontal Microbial Dark MatterDocument16 pagesPeriodontal Microbial Dark MatterAndrés Nicolás Beltrán SeguraNo ratings yet
- HIV A Virus Like No OtherDocument83 pagesHIV A Virus Like No Otherbadboy6.868510No ratings yet
- Dunn RefDocument47 pagesDunn RefFlorencia Chandrika HalimNo ratings yet
- (Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springer International Publishing - Humana Press (2019)Document406 pages(Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springer International Publishing - Humana Press (2019)karima majdouliNo ratings yet
- (Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springe PDFDocument406 pages(Current Cancer Research) Erle S. Robertson - Microbiome and Cancer-Springe PDFmariomgNo ratings yet
- What Is CancerDocument2 pagesWhat Is CancerDebapriya HazraNo ratings yet
- Human Pathology (2008) 39, 154-166Document13 pagesHuman Pathology (2008) 39, 154-166Laura valentina ramos mezaNo ratings yet
- First Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDocument11 pagesFirst Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDavid A. Osorio G.No ratings yet
- Origin and Cellular Evolution-Claudiu BandeaDocument16 pagesOrigin and Cellular Evolution-Claudiu BandeaMedicina Usmp Filial NorteNo ratings yet
- Viruses & EvolutionDocument27 pagesViruses & Evolutionhaluk bilginNo ratings yet
- Exosomes - The Protagonists in The Tale of Colorectal CancerDocument38 pagesExosomes - The Protagonists in The Tale of Colorectal CancerManoj Kumar pandreNo ratings yet
- Research Paper On SmallpoxDocument11 pagesResearch Paper On Smallpoxxsykcbikf100% (1)
- The Role of Human Papillomavirus in Pre-Cancerous Lesions and Oral CancersDocument29 pagesThe Role of Human Papillomavirus in Pre-Cancerous Lesions and Oral CancerssevattapillaiNo ratings yet
- Notes 1Document5 pagesNotes 1logonautNo ratings yet
- Chronic CancerDocument13 pagesChronic Cancerjulius_henryNo ratings yet
- Tropicalmed 08 00173 v2Document12 pagesTropicalmed 08 00173 v2cesar martin seguraNo ratings yet
- David S. Lindsay, Louis M. Weiss - Opportunistic Infections - Toxoplasma, Sarcocystis, and Microsporidia (World Class Parasites) (2004)Document249 pagesDavid S. Lindsay, Louis M. Weiss - Opportunistic Infections - Toxoplasma, Sarcocystis, and Microsporidia (World Class Parasites) (2004)DuONo ratings yet
- The Dna Provirus HypothesisDocument10 pagesThe Dna Provirus HypothesismarsbleachNo ratings yet
- Jackie Virology AssignmentDocument6 pagesJackie Virology AssignmentDumisani NguniNo ratings yet
- Infectious Disease Ecology of Wild Birds Jennifer C Owen Full ChapterDocument48 pagesInfectious Disease Ecology of Wild Birds Jennifer C Owen Full Chapterray.marks344100% (12)
- 10201148Document96 pages10201148Huzaifa SyedNo ratings yet
- Transformation and Oncogenesis: Biology W3310/4310 Virology Spring 2016Document56 pagesTransformation and Oncogenesis: Biology W3310/4310 Virology Spring 2016kashif manzoorNo ratings yet
- The Basics of VirusesDocument4 pagesThe Basics of VirusesSharolt SierraNo ratings yet
- From Mendel To The Human Genome Project: The Implications For Nurse EducationDocument6 pagesFrom Mendel To The Human Genome Project: The Implications For Nurse EducationKome ČemuNo ratings yet
- TOTUL Despre HPVDocument220 pagesTOTUL Despre HPVsweetchildofmineNo ratings yet
- Waqar AhmedDocument11 pagesWaqar AhmedSyed Hassan GillaniNo ratings yet
- Viruses: A Scientific American Library BookFrom EverandViruses: A Scientific American Library BookRating: 4.5 out of 5 stars4.5/5 (11)
- BT 2019Document13 pagesBT 2019biotech_vidhyaNo ratings yet
- Q.No. Type Section Key/Range MarksDocument3 pagesQ.No. Type Section Key/Range Marksbiotech_vidhyaNo ratings yet
- Troubleshooting SDS-PAGE 1Document3 pagesTroubleshooting SDS-PAGE 1biotech_vidhyaNo ratings yet
- Buffer Preparation PDFDocument6 pagesBuffer Preparation PDFbiotech_vidhyaNo ratings yet
- Whole Cell ExtractDocument1 pageWhole Cell Extractbiotech_vidhyaNo ratings yet
- Polymerase Chain Reaction (PCR)Document3 pagesPolymerase Chain Reaction (PCR)biotech_vidhyaNo ratings yet
- Nuclear ExtractsDocument2 pagesNuclear Extractsbiotech_vidhyaNo ratings yet
- Buffer Preparation PDFDocument6 pagesBuffer Preparation PDFbiotech_vidhyaNo ratings yet
- Ies 17 Set A Me Q ADocument67 pagesIes 17 Set A Me Q Abiotech_vidhyaNo ratings yet
- Mechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010Document20 pagesMechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010biotech_vidhyaNo ratings yet
- Befcv List PDFDocument22 pagesBefcv List PDFbiotech_vidhyaNo ratings yet
- TNPSC Group 1 Prelim Book List PDFDocument2 pagesTNPSC Group 1 Prelim Book List PDFbiotech_vidhyaNo ratings yet
- 1 TolerancesDocument1 page1 Tolerancesbiotech_vidhyaNo ratings yet
- Teachers Recruitment Board: 1. Important DatesDocument13 pagesTeachers Recruitment Board: 1. Important Datesbiotech_vidhyaNo ratings yet
- Isa BusDocument30 pagesIsa Busbiotech_vidhya100% (1)
- Lec09 Three PhaseDocument84 pagesLec09 Three Phasebiotech_vidhyaNo ratings yet
- K - 1 Perspective and The Future of OncologyDocument36 pagesK - 1 Perspective and The Future of OncologyPRAGAATHYRAJANNo ratings yet
- Imunologi Tumor: Syarifuddin WahidDocument40 pagesImunologi Tumor: Syarifuddin WahidinhaNo ratings yet
- Decoding Cancer - Lesson 1 - Interactive Lesson - July - 2018Document46 pagesDecoding Cancer - Lesson 1 - Interactive Lesson - July - 2018NisrinNo ratings yet
- Ch4 Lecture SlidesDocument34 pagesCh4 Lecture SlidesJad AwadNo ratings yet
- This Study Resource Was: G R A D E S L A B - C O MDocument8 pagesThis Study Resource Was: G R A D E S L A B - C O MHugsNo ratings yet
- 02 STR 9 102Document94 pages02 STR 9 10211111No ratings yet
- Neoplasia PDFDocument182 pagesNeoplasia PDFmarwaNo ratings yet
- Cancer Genetics: Sreekutty S 2 MSC ZoologyDocument34 pagesCancer Genetics: Sreekutty S 2 MSC ZoologyShamsudheen maharajasNo ratings yet
- Genomes and Their EvolutionDocument23 pagesGenomes and Their Evolutionprehealthhelp100% (2)
- Emperor of All Maladies CastDocument6 pagesEmperor of All Maladies Castmykwin07No ratings yet
- FMS 1 - Week 7 Tutorial 1 LO PDFDocument17 pagesFMS 1 - Week 7 Tutorial 1 LO PDFAprillia AlmaasNo ratings yet
- Plants That Fight CancerDocument314 pagesPlants That Fight CancerMai Elnaggar100% (4)
- Chapter 14 Mutation and CancerDocument43 pagesChapter 14 Mutation and CancerCharlieHuangNo ratings yet
- Neoplasia: Nomenclature Epidemiology OncogenesDocument7 pagesNeoplasia: Nomenclature Epidemiology OncogenespriyaNo ratings yet
- Oncogene EtcDocument48 pagesOncogene EtcInsyirah JohariNo ratings yet
- General Biology 1: Quarter 1 - Module - : Title: Cell CycleDocument27 pagesGeneral Biology 1: Quarter 1 - Module - : Title: Cell CycleRea A. Bilan0% (1)
- CarcinogenesisDocument3 pagesCarcinogenesiskishorkumarn8212No ratings yet
- Cytogenetics Final ExamDocument20 pagesCytogenetics Final ExamMICHAEL JOHN AGUILARNo ratings yet
- 6 Hos CarcinogenesisDocument90 pages6 Hos CarcinogenesisalgutNo ratings yet
- NikitaDocument41 pagesNikitaAtika AnggrainiNo ratings yet
- 1.biokimia KankerDocument94 pages1.biokimia Kankersebastian chendraNo ratings yet
- Onco, TSG & CancerDocument8 pagesOnco, TSG & Cancersumera120488No ratings yet
- Chemotherapy of Cancer: Submitted by Syama - J.S Roll No:15 2 SEM M Pharm PharmacologyDocument64 pagesChemotherapy of Cancer: Submitted by Syama - J.S Roll No:15 2 SEM M Pharm PharmacologySyama J.S100% (1)
- Quiz BankDocument12 pagesQuiz Banknadim1241335No ratings yet
- Tumor VirusesDocument28 pagesTumor VirusespavindersinghNo ratings yet