Download as pdf or txt
You might also like
- Unit 5 Individual Project 1Document6 pagesUnit 5 Individual Project 1MUNSIF JAWEEDNo ratings yet
- MQM100 MultipleChoice Chapter3Document21 pagesMQM100 MultipleChoice Chapter3Nakin K100% (2)
- 96 ReptilelifehistoryDocument13 pages96 ReptilelifehistoryRebeccaSulivanNo ratings yet
- How Old Is Human LongevityDocument4 pagesHow Old Is Human LongevityRoberto Oscar LandaNo ratings yet
- Newton Et Al 2016 Raptor Survival Review Ibis PDFDocument22 pagesNewton Et Al 2016 Raptor Survival Review Ibis PDFmariaNo ratings yet
- K Development Team: Zoology Principles of Ecology Life Table, Fecundity Table and Survivorship Curves, Part-IDocument17 pagesK Development Team: Zoology Principles of Ecology Life Table, Fecundity Table and Survivorship Curves, Part-IIlham ArfiansyahNo ratings yet
- Sergio Et Al 2011 Milvus Migrans Demography Age StructuredDocument9 pagesSergio Et Al 2011 Milvus Migrans Demography Age StructuredmariaNo ratings yet
- Age-At-Death Standards For Mesoamerican Prehispanic and Colonial Infant, Child, and Juvenile SkeletonsDocument18 pagesAge-At-Death Standards For Mesoamerican Prehispanic and Colonial Infant, Child, and Juvenile SkeletonsLuisa Fernanda GonzalezNo ratings yet
- Harvey & Clutton - Life History Variation in PrimatesDocument23 pagesHarvey & Clutton - Life History Variation in PrimatesCarlos PlazasNo ratings yet
- Idade Elefante 2Document10 pagesIdade Elefante 2Fernanda MatosNo ratings yet
- 2011 RobbinsDocument6 pages2011 Robbinsgwen_schugNo ratings yet
- Roksandic y Armstrong (2011)Document11 pagesRoksandic y Armstrong (2011)silviahuesosNo ratings yet
- 52a 3 B171 PDFDocument8 pages52a 3 B171 PDFManasaNo ratings yet
- Seasonal Mortality Patterns in Non-Human Primates: Implications For Variation in Selection Pressures Across EnvironmentsDocument15 pagesSeasonal Mortality Patterns in Non-Human Primates: Implications For Variation in Selection Pressures Across Environmentsapi-527444510No ratings yet
- Ontogeny of Skull Size and Shape Changes Within A Framework of Biphasic Lifestyle: A Case Study in Six Triturus Species (Amphibia, Salamandridae)Document11 pagesOntogeny of Skull Size and Shape Changes Within A Framework of Biphasic Lifestyle: A Case Study in Six Triturus Species (Amphibia, Salamandridae)mikimaricNo ratings yet
- 2003P v32n2p327Document7 pages2003P v32n2p327Mehmet Salih ÖzgökçeNo ratings yet
- 2003P v32n2p327Document7 pages2003P v32n2p327Mehmet Salih ÖzgökçeNo ratings yet
- Genetic Variation and Polymorphim in The Inducible Spines of A Marine BryozoanDocument13 pagesGenetic Variation and Polymorphim in The Inducible Spines of A Marine BryozoanCarlos MeirellesNo ratings yet
- Evo 13845Document12 pagesEvo 13845ELLENA SANTOSNo ratings yet
- JAS InfantDocument18 pagesJAS Infantanni rahimaNo ratings yet
- 2008 GreenfieldandArnold Sheep Goat Tooth Wear Eruption AgeDocument15 pages2008 GreenfieldandArnold Sheep Goat Tooth Wear Eruption AgeSidra ZahoorNo ratings yet
- Invariantes Biologicos 1Document5 pagesInvariantes Biologicos 1Paulo Pedro P. R. CostaNo ratings yet
- Longevity Among Hunter-GatherersDocument46 pagesLongevity Among Hunter-GatherersTomasz IgnasiakNo ratings yet
- Population Density, Survival, Growth, and Development of The American CockroachDocument18 pagesPopulation Density, Survival, Growth, and Development of The American CockroachjohnNo ratings yet
- Kura Ku 2008Document13 pagesKura Ku 2008J.D. NobleNo ratings yet
- Litter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Document12 pagesLitter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Tayssa MarquesNo ratings yet
- Effect of Age and Parity On Litter Size and OffsprDocument7 pagesEffect of Age and Parity On Litter Size and OffsprAulia Fauziyyah RahmiNo ratings yet
- Journal EmbrioDocument9 pagesJournal Embrio'Immey Ssi MoyNo ratings yet
- Costa-Souza Et Al 2019 - Growth, Sexual Maturity and Dimorphism in Six Species of SnappingDocument14 pagesCosta-Souza Et Al 2019 - Growth, Sexual Maturity and Dimorphism in Six Species of Snappingkeltony aquinoNo ratings yet
- Raziyeh Alaei Life History Traits and The FirstDocument16 pagesRaziyeh Alaei Life History Traits and The FirstalaeiraziyeNo ratings yet
- Shine, 1990Document7 pagesShine, 1990Leticia MioloNo ratings yet
- Seasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientDocument21 pagesSeasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientVíctor RamirezdNo ratings yet
- Abegg Et Al., 2020Document10 pagesAbegg Et Al., 2020Míriam DuarteNo ratings yet
- Gallardo Et Al 2006 (Kingiella Chilenica)Document10 pagesGallardo Et Al 2006 (Kingiella Chilenica)j.salazar.somNo ratings yet
- 1 s2.0 S0032579119353490 MainDocument8 pages1 s2.0 S0032579119353490 MainTerence John AguinaldoNo ratings yet
- E Meind98Document25 pagesE Meind98Sisesti Primaria SisestiNo ratings yet
- Linares Et Al. 2007 EcologyDocument11 pagesLinares Et Al. 2007 EcologyFadhilla SetyaNo ratings yet
- Formal Report 7Document8 pagesFormal Report 7PAUL CEDRIC BERNARDONo ratings yet
- T Z N - Et - Al 2018 OikosDocument11 pagesT Z N - Et - Al 2018 OikosSuany Quesada CalderonNo ratings yet
- Applied Animal Ethology, Campitelli1982Document8 pagesApplied Animal Ethology, Campitelli1982Mohamed khNo ratings yet
- Elephant SealDocument9 pagesElephant SealAbraham MartinezNo ratings yet
- Short-Term e Ffects of Fishing On Life History Traits of FishesDocument21 pagesShort-Term e Ffects of Fishing On Life History Traits of FishesssotomNo ratings yet
- The Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Document9 pagesThe Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Евгений ЯворскийNo ratings yet
- Geometric Morphometric Analysis of Body-Form Variability In: Cichlasoma Minckleyi, The Cuatro Cienegas CichlidDocument13 pagesGeometric Morphometric Analysis of Body-Form Variability In: Cichlasoma Minckleyi, The Cuatro Cienegas CichlidArben Anthony Saavedra QuitosNo ratings yet
- Longevity Among Hunter-Gatherers: A Cross-Cultural ExaminationDocument45 pagesLongevity Among Hunter-Gatherers: A Cross-Cultural ExaminationYamil SaieghNo ratings yet
- Baptista Et Al. 2000. Postnatal Growth and Age Estimation in Free-Ranging Bats A Comparison of Longitudinal and Cross-Sectional Sampling Methods PDFDocument10 pagesBaptista Et Al. 2000. Postnatal Growth and Age Estimation in Free-Ranging Bats A Comparison of Longitudinal and Cross-Sectional Sampling Methods PDFHenry CondoriNo ratings yet
- Dermochelys Coriacea SkeletonDocument13 pagesDermochelys Coriacea SkeletonYessica VargasNo ratings yet
- KamelOyarzunGrosberg 2010Document12 pagesKamelOyarzunGrosberg 2010fygv1990No ratings yet
- End of Biological ReproductionDocument1 pageEnd of Biological ReproductionCadell LastNo ratings yet
- Artigo 7 - Biotic Potencial and Reproductive Parameters of Spodoptera Eridania (Stoll) (Lepidoptera, Noctuidae) in The LaboratoryDocument7 pagesArtigo 7 - Biotic Potencial and Reproductive Parameters of Spodoptera Eridania (Stoll) (Lepidoptera, Noctuidae) in The Laboratoryjean alex thimotheeNo ratings yet
- Bar-Oz y Munro 2006Document11 pagesBar-Oz y Munro 2006Ana SbattellaNo ratings yet
- Alexandre Specht - Agrotis Malefida - ROUND 1 Revisado Pelo Autor - JEFFDocument22 pagesAlexandre Specht - Agrotis Malefida - ROUND 1 Revisado Pelo Autor - JEFFsamusaran115No ratings yet
- Sex-Specific Seasonal Body Mass Variation in The Pale-Breasted Thrush (Turdus Leucomelas)Document5 pagesSex-Specific Seasonal Body Mass Variation in The Pale-Breasted Thrush (Turdus Leucomelas)Jani PereiraNo ratings yet
- 05 Chroma LV JsemDocument10 pages05 Chroma LV JsemMaryzender Rodríguez MelgarejoNo ratings yet
- TMP E39Document41 pagesTMP E39FrontiersNo ratings yet
- Life Tables, Survivorship Curves & Age-Sex Structure (Article) - Khan Academy PDFDocument20 pagesLife Tables, Survivorship Curves & Age-Sex Structure (Article) - Khan Academy PDFIrish MercadoNo ratings yet
- GrantGrant2000 PDFDocument9 pagesGrantGrant2000 PDFahmed abdulatifNo ratings yet
- FulltextDocument4 pagesFulltextMohammed KouidriNo ratings yet
- Johnson 2000Document26 pagesJohnson 2000IASA73No ratings yet
- MC KayDocument4 pagesMC Kayanamia28No ratings yet
- Eviscerated Yield, Component Parts, and Meat, Skin and Bone Ratios in The Chicken Broiler"Document5 pagesEviscerated Yield, Component Parts, and Meat, Skin and Bone Ratios in The Chicken Broiler"Mahbub RezaNo ratings yet
- Key to the identification and ecology of Cyclopoida (Crustacea, Copepoda) of North America (north of Mexico)From EverandKey to the identification and ecology of Cyclopoida (Crustacea, Copepoda) of North America (north of Mexico)No ratings yet
- Demand Forecasting LectureDocument68 pagesDemand Forecasting Lectureapi-26872269100% (3)
- Dragon Quest 8 Skills ListDocument10 pagesDragon Quest 8 Skills ListsorwenNo ratings yet
- S1 Edexcel Revision PackDocument9 pagesS1 Edexcel Revision PackJFGHANSAHNo ratings yet
- W 10 Markov ModelDocument13 pagesW 10 Markov ModelLawlietNo ratings yet
- Significant FiguresDocument5 pagesSignificant FiguresAradhana YadavNo ratings yet
- Minitab Multiple Regression AnalysisDocument6 pagesMinitab Multiple Regression AnalysisEka Rahma Paramita100% (1)
- Mutli Component DistillationDocument13 pagesMutli Component Distillationramantal5No ratings yet
- CH 123Document63 pagesCH 123Talha ImtiazNo ratings yet
- Persona 4 WalkthroughDocument194 pagesPersona 4 WalkthroughAtenk BadguyNo ratings yet
- Reduction of Concrete Sorptivity With Age Through CarbonationDocument7 pagesReduction of Concrete Sorptivity With Age Through CarbonationSritam SwapnadarshiNo ratings yet
- Managerial Economics (Quiz) Subject Code: 12MCEC01 MBA-Trimester - IDocument4 pagesManagerial Economics (Quiz) Subject Code: 12MCEC01 MBA-Trimester - IShivani PandeyNo ratings yet
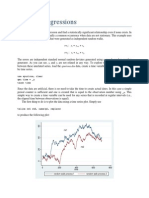
- Spurious Regressions: RW y y V RW X X VDocument3 pagesSpurious Regressions: RW y y V RW X X Vkaddour7108No ratings yet
- D7bc1home Assignment 1Document5 pagesD7bc1home Assignment 1Mayank MittalNo ratings yet
- Revised Pavement Design - Beawar Pali Section PDFDocument119 pagesRevised Pavement Design - Beawar Pali Section PDFSanket ShahNo ratings yet
- Horizontal Distance MeasurementDocument24 pagesHorizontal Distance MeasurementAngelinne Catalan0% (1)
- Market AppraisalDocument19 pagesMarket AppraisalCharu SharmaNo ratings yet
- Estimating Risk and Return On Assets: S A R Q P I. QuestionsDocument13 pagesEstimating Risk and Return On Assets: S A R Q P I. QuestionsRonieOlarteNo ratings yet
- Time Series Analysis Exercises: Universität PotsdamDocument30 pagesTime Series Analysis Exercises: Universität PotsdamSD100% (1)
- CLIMDEX: Climate Extremes IndicesDocument5 pagesCLIMDEX: Climate Extremes Indicessuraj.atmos458No ratings yet
- Problem IDocument10 pagesProblem Imgharba123No ratings yet
- Multistage SamplingDocument29 pagesMultistage SamplingRizki Amalia Puji SantosaNo ratings yet
- Double-Mass Curve USGSDocument41 pagesDouble-Mass Curve USGSrajesh005100% (1)
- Erf (T)Document44 pagesErf (T)brayan2904No ratings yet
- Ixl Math Common Core Alignments 6Document13 pagesIxl Math Common Core Alignments 6api-293959403No ratings yet
- AS Manual PDFDocument76 pagesAS Manual PDFPurwanto NugrohoNo ratings yet
- Ian Wallace - Introduction To Project Risk Management Using @RISK For ProjectDocument12 pagesIan Wallace - Introduction To Project Risk Management Using @RISK For ProjectAhmed RaufNo ratings yet
- Stochastic Integration With Jumps (Bichteler)Document645 pagesStochastic Integration With Jumps (Bichteler)dsussex123No ratings yet
- 0-10 CMC CalculationDocument4 pages0-10 CMC CalculationAgus TNo ratings yet