Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Community Process, Dynamics & EmpowermentDocument30 pagesCommunity Process, Dynamics & EmpowermentRateliaNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Readworks - From One To ManyDocument5 pagesReadworks - From One To ManyPilar Andrea MarquezNo ratings yet
- Cosmetology Portfolio: Jaimie R. FortinDocument34 pagesCosmetology Portfolio: Jaimie R. FortinJaimie FantasticNo ratings yet
- Interpersonel Need of Management Student-Acilitor in The Choice of ElectivesDocument180 pagesInterpersonel Need of Management Student-Acilitor in The Choice of ElectivesnerdjumboNo ratings yet
- Project SideshowDocument4 pagesProject SideshowMetro English CanadaNo ratings yet
- The Institute of Chartered Secretaries of Bangladesh (ICSB) : FCS, Fca, Fcma, Mba, LL.BDocument8 pagesThe Institute of Chartered Secretaries of Bangladesh (ICSB) : FCS, Fca, Fcma, Mba, LL.BLubainur RahmanNo ratings yet
- Pplication FOR Mployment: (Please Do Not Change Font Style or Font Size When You Fill Out The Form)Document4 pagesPplication FOR Mployment: (Please Do Not Change Font Style or Font Size When You Fill Out The Form)Lubainur RahmanNo ratings yet
- Assignment Cover Sheet: For Submission of Individual and Group WorkDocument1 pageAssignment Cover Sheet: For Submission of Individual and Group WorkLubainur RahmanNo ratings yet
- The Tragedy of The CommonsDocument1 pageThe Tragedy of The CommonsLubainur RahmanNo ratings yet
- Page 3Document1 pagePage 3Lubainur RahmanNo ratings yet
- Assessing The Brand Awareness of Consumers On Boutique and Fashion Houses in Bangladesh - A Study On AarongDocument5 pagesAssessing The Brand Awareness of Consumers On Boutique and Fashion Houses in Bangladesh - A Study On AarongLubainur RahmanNo ratings yet
- DefinitionDocument2 pagesDefinitionLubainur RahmanNo ratings yet
- Page 4Document1 pagePage 4Lubainur RahmanNo ratings yet
- Post Transcriptional ModificationDocument3 pagesPost Transcriptional ModificationLubainur RahmanNo ratings yet
- Brinjal BreedingDocument25 pagesBrinjal BreedingLubainur RahmanNo ratings yet
- Shahin CVDocument2 pagesShahin CVLubainur RahmanNo ratings yet
- Criminal SociologyDocument66 pagesCriminal SociologySal McGaundieNo ratings yet
- Instruction Manual TW25 (9240Y) POWERPOINT-DishwaherDocument20 pagesInstruction Manual TW25 (9240Y) POWERPOINT-DishwahergreengawNo ratings yet
- CookiesDocument3 pagesCookiesjonajonajonaNo ratings yet
- Previous HSE Questions From The Chapter "ELECTROCHEMISTRY": E E - 2.303RT Log (M) NF (M)Document2 pagesPrevious HSE Questions From The Chapter "ELECTROCHEMISTRY": E E - 2.303RT Log (M) NF (M)Chemistry MESNo ratings yet
- Ujian Setara 1 2017 KimiaDocument9 pagesUjian Setara 1 2017 KimiaZulkifli Bin PariNo ratings yet
- B1 Questions QN001: Form Questions Which Ask For The Underlined WordsDocument2 pagesB1 Questions QN001: Form Questions Which Ask For The Underlined WordsMirjana Paunovic100% (1)
- A Case Study of The Dublin and Galway Rape Crisis Centres: The Therapist'S PerspectiveDocument196 pagesA Case Study of The Dublin and Galway Rape Crisis Centres: The Therapist'S PerspectiveAG VenturesNo ratings yet
- Excavation and Shoring SH-2012-23eDocument5 pagesExcavation and Shoring SH-2012-23eshamroz khan100% (1)
- Sample QuestionnaireDocument5 pagesSample QuestionnaireAudi Kyle SaydovenNo ratings yet
- Author Guidline. CPSPDocument4 pagesAuthor Guidline. CPSPMuhammad MoazzamNo ratings yet
- Arizona Communicable Disease FlipchartDocument98 pagesArizona Communicable Disease Flipchartapi-510866696No ratings yet
- Training Manual For Gender TrainersDocument43 pagesTraining Manual For Gender TrainersMiqz ZenNo ratings yet
- Racial Discrimination: Examples For RacismDocument4 pagesRacial Discrimination: Examples For RacismGopika MoorkothNo ratings yet
- Butternut Squash SoupDocument1 pageButternut Squash SouplyllacycusNo ratings yet
- Drug Education and Vice ControlDocument16 pagesDrug Education and Vice ControlSinagTalaNo ratings yet
- 5800 Om PDFDocument104 pages5800 Om PDFPré ImpressãoNo ratings yet
- Coca Cola Peruvian EconomicsDocument34 pagesCoca Cola Peruvian Economicsborn2dive 9702No ratings yet
- Review: Recent Approaches in Herbal Drug StandardizationDocument9 pagesReview: Recent Approaches in Herbal Drug Standardizationkj0203No ratings yet
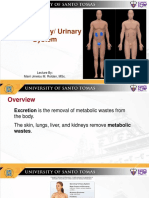
- The Excretory/ Urinary System: Lecture By: Marri Jmelou M. Roldan, MSCDocument29 pagesThe Excretory/ Urinary System: Lecture By: Marri Jmelou M. Roldan, MSCHeaven SolNo ratings yet
- Price List: Heavy Duty Foodwaste Disposers and Electrical ControlsDocument39 pagesPrice List: Heavy Duty Foodwaste Disposers and Electrical Controlsedgar ricoNo ratings yet
- Ultra High Temperature 100HTL Adhesive Transfer Tapes: Technical Data November, 2006Document4 pagesUltra High Temperature 100HTL Adhesive Transfer Tapes: Technical Data November, 2006Iris SoonNo ratings yet
- Research Proposal - Matthew WengrowichDocument17 pagesResearch Proposal - Matthew Wengrowichapi-311840545100% (1)
- ELECTROMYOGRAPHYDocument7 pagesELECTROMYOGRAPHYsariNo ratings yet
- Ridf.Document21 pagesRidf.Syam KrishnanNo ratings yet
- Nabl 400Document669 pagesNabl 400Apex IndiaNo ratings yet