Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Shortness of Breath: Checklist PMPF Checklist PMPFDocument1 pageShortness of Breath: Checklist PMPF Checklist PMPFanz_4191No ratings yet
- PDFDocument422 pagesPDFThuRein WinNo ratings yet
- Benefits of Gene TechnologyDocument29 pagesBenefits of Gene TechnologyAngga Sendiputra100% (1)
- The Value of Negative EmotionsDocument3 pagesThe Value of Negative EmotionsFred P. GalloNo ratings yet
- Occupational English Test: Writing Subtest: Nursing Time Allowed: Reading Time: 5 Minutes Writing Time: 40 MinutesDocument2 pagesOccupational English Test: Writing Subtest: Nursing Time Allowed: Reading Time: 5 Minutes Writing Time: 40 MinutesHarold MedranoNo ratings yet
- Commonwealth of Australia Warning: Do Not Remove This NoticeDocument40 pagesCommonwealth of Australia Warning: Do Not Remove This NoticeDerek ChoyNo ratings yet
- Hon 2005Document5 pagesHon 2005lenirizkiNo ratings yet
- Anisometropia 4Document18 pagesAnisometropia 4Mamtha MMNo ratings yet
- Dimeglio 2012Document12 pagesDimeglio 2012Tantia Dewi HariantoNo ratings yet
- Nursing Care in ECTDocument3 pagesNursing Care in ECTRawan KhateebNo ratings yet
- Surya Herbals PDFDocument5 pagesSurya Herbals PDFVenkatesh MurthyNo ratings yet
- Dinaburg Eval 2Document1 pageDinaburg Eval 2api-354606946No ratings yet
- Ethnobotanical AwarenessDocument7 pagesEthnobotanical AwarenessFranzine L.No ratings yet
- Zinc OxideDocument4 pagesZinc OxideyundayundaaNo ratings yet
- 13 Alvarez II vs. Sun Life of CanadaDocument1 page13 Alvarez II vs. Sun Life of CanadaPaolo AlarillaNo ratings yet
- (Bioethics 2) 2.01.01 - Case - Karen Quinlan (Pat G)Document1 page(Bioethics 2) 2.01.01 - Case - Karen Quinlan (Pat G)Mikee MeladNo ratings yet
- The Antibacterial Efficacy of Gotu Kola (Centella Asiatica) - Base SoapDocument11 pagesThe Antibacterial Efficacy of Gotu Kola (Centella Asiatica) - Base SoapJohnnard BelenNo ratings yet
- Single Subject ResearchDocument24 pagesSingle Subject ResearchYogi Saputra Mahmud100% (1)
- AOS ERP Generic FlowchartDocument4 pagesAOS ERP Generic FlowchartRidha JuliyantiNo ratings yet
- Circular Cghs Referal Expostfacto Approval Delegation 7-4-99Document4 pagesCircular Cghs Referal Expostfacto Approval Delegation 7-4-99Prakash Kapoor50% (2)
- SignagesDocument4 pagesSignagesRahul NegiNo ratings yet
- Soal PAS B.Ing Kelas 9Document5 pagesSoal PAS B.Ing Kelas 9DidikSakLaekEtohNo ratings yet
- Army Public School Basistha InvestigatorDocument25 pagesArmy Public School Basistha InvestigatorSuresh Kumar SubbiahNo ratings yet
- Teaching StrategiesDocument15 pagesTeaching StrategiesJuliansyaNo ratings yet
- Senarai Nama Murid RBT Tahun 4Document5 pagesSenarai Nama Murid RBT Tahun 4aziziishak100% (1)
- Class Record: Students Name Attendance QuizzesDocument4 pagesClass Record: Students Name Attendance QuizzesKarl Angelo MontanoNo ratings yet
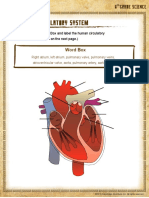
- Label The Circulatory SystemDocument2 pagesLabel The Circulatory SystemIzzu PenielNo ratings yet
- Test Bank For Ecgs Made Easy 4th Edition Barbara J AehlertDocument12 pagesTest Bank For Ecgs Made Easy 4th Edition Barbara J AehlertRonald Herron100% (38)
- Runers Do It Better - Sex!Document2 pagesRuners Do It Better - Sex!webizerNo ratings yet
- July, 2023 Nursing and Training Report.Document20 pagesJuly, 2023 Nursing and Training Report.Tosi AboloNo ratings yet