Download as pdf or txt
You might also like
- Cell Membrane 100 OnlineDocument38 pagesCell Membrane 100 OnlineNur Ayunni KhafizNo ratings yet
- Met Sy & Circadian RyhthmDocument3 pagesMet Sy & Circadian RyhthmSameh Ali ElsayedNo ratings yet
- Therapeutic Investigation of Galic Acid On Paclitaxel-Induced Motor-Incoordination in Mus MusculusDocument5 pagesTherapeutic Investigation of Galic Acid On Paclitaxel-Induced Motor-Incoordination in Mus MusculusRAPPORTS DE PHARMACIENo ratings yet
- Evidence For A Role of KISS-1/GPR54 System in Decreased Luteinizing Hormone (LH) Secretion in Fasted Prepubertal EwesDocument7 pagesEvidence For A Role of KISS-1/GPR54 System in Decreased Luteinizing Hormone (LH) Secretion in Fasted Prepubertal EwesAndrea CelisNo ratings yet
- Article - Metabolism in Rats During Antiorthostatic HypokinesiaDocument3 pagesArticle - Metabolism in Rats During Antiorthostatic HypokinesianastiaNo ratings yet
- In Vivo: Xanthine Oxidase Inhibition StudiesDocument16 pagesIn Vivo: Xanthine Oxidase Inhibition StudiesHartono TanambellNo ratings yet
- Physiological Effects of Melatonin On Leptin, Testosterone and Biochemical Parameters in Albino RatsDocument6 pagesPhysiological Effects of Melatonin On Leptin, Testosterone and Biochemical Parameters in Albino RatsIOSR Journal of PharmacyNo ratings yet
- Uludag 2007 Twentyfourhour Variation OflargininDocument13 pagesUludag 2007 Twentyfourhour Variation OflargininNurettin AbacıoğluNo ratings yet
- Huitron-Resendiz 2001 PDFDocument40 pagesHuitron-Resendiz 2001 PDFМатиас Себальос ГусманNo ratings yet
- Sun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDocument7 pagesSun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDaniela Patricia Alvarez AravenaNo ratings yet
- Effects of Magnesium On Cytomorphology and Enzyme Activities in Thyroids of RatsDocument6 pagesEffects of Magnesium On Cytomorphology and Enzyme Activities in Thyroids of RatsPallav SenguptaNo ratings yet
- 1414 431X BJMBR 1414 431X20176346Document8 pages1414 431X BJMBR 1414 431X20176346Guilherme Vasconcelos VasconcelosNo ratings yet
- Strychnine: (CAS No: 57-24-9) Health-Based Reassessment of Administrative Occupational Exposure LimitsDocument20 pagesStrychnine: (CAS No: 57-24-9) Health-Based Reassessment of Administrative Occupational Exposure LimitsDhira GunawanNo ratings yet
- Chair Restraint in MonkeyDocument7 pagesChair Restraint in MonkeyveganjermNo ratings yet
- 28sici 291097 4547 2819961101 2946 3A3 3C316 3A 3aaid jnr4 3e3.0.co 3B2 NDocument8 pages28sici 291097 4547 2819961101 2946 3A3 3C316 3A 3aaid jnr4 3e3.0.co 3B2 NLata DeshmukhNo ratings yet
- R. E. PertiwiDocument13 pagesR. E. PertiwiNickoNo ratings yet
- 407 09 05 ArticleDocument13 pages407 09 05 ArticleAnna ShofiyaNo ratings yet
- The Effect of Monosodium Glutamate On Blood Glucose Levels in MiceDocument9 pagesThe Effect of Monosodium Glutamate On Blood Glucose Levels in Micepratiwi kusumaNo ratings yet
- Pier Vincenzo Piazza Et Al - Glucocorticoids Have State-Dependent Stimulant Effects On The Mesencephalic Dopaminergic TransmissionDocument5 pagesPier Vincenzo Piazza Et Al - Glucocorticoids Have State-Dependent Stimulant Effects On The Mesencephalic Dopaminergic TransmissionSour60No ratings yet
- Sandip R Final SynopsisDocument15 pagesSandip R Final SynopsisnikuNo ratings yet
- Tugas Toksikologi Parasetamol Dina Yuspita Sari (Nim h2061151002)Document30 pagesTugas Toksikologi Parasetamol Dina Yuspita Sari (Nim h2061151002)putryNo ratings yet
- NMDA/glutamate Mechanism of Anxiolytic-Like Action of Zinc in Vogel Conflict Test in MiceDocument9 pagesNMDA/glutamate Mechanism of Anxiolytic-Like Action of Zinc in Vogel Conflict Test in MiceIOSR Journal of PharmacyNo ratings yet
- Fuentealba J Et Al 2007Document4 pagesFuentealba J Et Al 2007Claudia Perez ManriquezNo ratings yet
- Thyroid Hormone SummaryDocument11 pagesThyroid Hormone SummaryAishwarya SinghNo ratings yet
- Trepan I Er 1991Document6 pagesTrepan I Er 1991Luisa Fernanda Rojas DiazNo ratings yet
- Cocaine and HipocampoDocument7 pagesCocaine and HipocampoNico TánatosNo ratings yet
- Effects of Dietary Magnesium On Testicular Histology, Steroidogenesis, Spermatogenesis and Oxidative Stress Markers in Adult RatsDocument11 pagesEffects of Dietary Magnesium On Testicular Histology, Steroidogenesis, Spermatogenesis and Oxidative Stress Markers in Adult RatsPallav SenguptaNo ratings yet
- Effects of Acepromazine and Midazolam Along With Ketamin On Haematological Parameters in YakDocument4 pagesEffects of Acepromazine and Midazolam Along With Ketamin On Haematological Parameters in YakTJPRC PublicationsNo ratings yet
- The Open Natural Products Journal, 2009, 2, 68-70Document3 pagesThe Open Natural Products Journal, 2009, 2, 68-70Valentina AnutaNo ratings yet
- Effect of Conditioned Fear Stress On Serotonin Metabolism in The Rat BrainDocument4 pagesEffect of Conditioned Fear Stress On Serotonin Metabolism in The Rat BrainJef_8No ratings yet
- 2020 Pathophysiological Changes in Female Rats With Estrous Cycle Disorder Induced by Long-Term Heat StressDocument10 pages2020 Pathophysiological Changes in Female Rats With Estrous Cycle Disorder Induced by Long-Term Heat StressemmanouelaapostolopoulouNo ratings yet
- Ikegami+et+al Endocrinology Vol156 p647-659Document13 pagesIkegami+et+al Endocrinology Vol156 p647-659sunNo ratings yet
- Lorcaserin Summary by 4th GroupDocument3 pagesLorcaserin Summary by 4th GroupdimbrutNo ratings yet
- Aebi CatalaseDocument8 pagesAebi CatalasehijazNo ratings yet
- Aai of CallistemonDocument3 pagesAai of CallistemonSachin PanditNo ratings yet
- Effect of Ten Beta-Adrenoceptor Blocking Agents On Spontaneous MotilityDocument6 pagesEffect of Ten Beta-Adrenoceptor Blocking Agents On Spontaneous MotilityIoana AntonesiNo ratings yet
- 1980 Marden InvolvementDocument8 pages1980 Marden Involvementspanners 12No ratings yet
- Hum. Reprod.-2009-Gordon-2618-28Document11 pagesHum. Reprod.-2009-Gordon-2618-28Roberto OrellanaNo ratings yet
- Gutman Et Al 2011cDocument4 pagesGutman Et Al 2011cRoee GutmanNo ratings yet
- Differential Susceptibility of BALB/c, C57BL/6N, and CF1 Mice To Photoperiod ChangesDocument6 pagesDifferential Susceptibility of BALB/c, C57BL/6N, and CF1 Mice To Photoperiod ChangesGabrielSteffenNo ratings yet
- Diabetes PDFDocument12 pagesDiabetes PDFDANIELA MALAGÓN MONTAÑONo ratings yet
- tmpCD0 TMPDocument5 pagestmpCD0 TMPFrontiersNo ratings yet
- Fitoterapia: Jianfeng Xue, Wenjun Ding, Yan LiuDocument5 pagesFitoterapia: Jianfeng Xue, Wenjun Ding, Yan LiuAamir SohailNo ratings yet
- Selective Impairment of Insulin Signalling in The Hypothalamus of Obese Zucker RatsDocument12 pagesSelective Impairment of Insulin Signalling in The Hypothalamus of Obese Zucker RatsKenjiOgoshiNo ratings yet
- Fitoterapia: Jianfeng Xue, Wenjun Ding, Yan LiuDocument5 pagesFitoterapia: Jianfeng Xue, Wenjun Ding, Yan LiuAamir SohailNo ratings yet
- 2457 4735 1 SMDocument8 pages2457 4735 1 SMAgus Prihanto PurnomoNo ratings yet
- Ref 4Document7 pagesRef 4Lateecka R KulkarniNo ratings yet
- Retinoid X Receptor Agonists Have Anti-Obesity Effects and Improve Insulin Sensitivity in Zucker Fa Fa RatsDocument8 pagesRetinoid X Receptor Agonists Have Anti-Obesity Effects and Improve Insulin Sensitivity in Zucker Fa Fa RatsKurnia Fitri AprillianaNo ratings yet
- Effect of Mast Cell Stabilizer KetotifenDocument9 pagesEffect of Mast Cell Stabilizer KetotifenEvelinaNo ratings yet
- Vivo Analyses of Experimental Animals (6,7)Document23 pagesVivo Analyses of Experimental Animals (6,7)aakash sahaNo ratings yet
- Pharmaco KineticsDocument4 pagesPharmaco Kineticsai aayNo ratings yet
- Rat Study 1Document7 pagesRat Study 1DRx Pradeep KumarNo ratings yet
- Ijmrhs Vol 1 Issue 1Document257 pagesIjmrhs Vol 1 Issue 1editorijmrhsNo ratings yet
- 1anil EtalDocument6 pages1anil EtaleditorijmrhsNo ratings yet
- Tmp8a04 TMPDocument13 pagesTmp8a04 TMPFrontiersNo ratings yet
- Alterations in Peripheral Purinergic and Muscarinic Signaling of Rat Bladder After Long-Term Fructose-Induced Metabolic SyndromeDocument13 pagesAlterations in Peripheral Purinergic and Muscarinic Signaling of Rat Bladder After Long-Term Fructose-Induced Metabolic SyndromeThalia KarampasiNo ratings yet
- Effect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsDocument8 pagesEffect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsBrian ColborneNo ratings yet
- Glutamato Desregulado É o Principal Fator de Estresse Das Doenças Psiquiátricas, Seu Precursor Glutamina Modula o GlutamatoDocument9 pagesGlutamato Desregulado É o Principal Fator de Estresse Das Doenças Psiquiátricas, Seu Precursor Glutamina Modula o Glutamatorubem.clementeNo ratings yet
- Chem. Senses 32: 41-49, 2007Document9 pagesChem. Senses 32: 41-49, 2007gas_yoe2No ratings yet
- Sato M Et Al. The Role of The Endocrine System in Feeding-Induced Tissue-Specific Circadian Entrainment. Cell Report July 2014Document16 pagesSato M Et Al. The Role of The Endocrine System in Feeding-Induced Tissue-Specific Circadian Entrainment. Cell Report July 2014Jason ColeNo ratings yet
- The Toxicity of Caffein: An experimental study on different species of animalsFrom EverandThe Toxicity of Caffein: An experimental study on different species of animalsNo ratings yet
- Jain School of Allied HealthDocument38 pagesJain School of Allied HealthBinu GeorgeNo ratings yet
- FBS Lab - BroucherDocument9 pagesFBS Lab - BroucherBinu GeorgeNo ratings yet
- AnatomyDocument4 pagesAnatomyBinu GeorgeNo ratings yet
- Mode of DNA Replication - Meselson-Stahl Experiment (Article) - Khan AcademyDocument18 pagesMode of DNA Replication - Meselson-Stahl Experiment (Article) - Khan AcademyBinu GeorgeNo ratings yet
- InstructionsDocument11 pagesInstructionsBinu GeorgeNo ratings yet
- Nutritional Disease - Britannica Online EncyclopediaDocument13 pagesNutritional Disease - Britannica Online EncyclopediaBinu GeorgeNo ratings yet
- Acid Phosphatase - Wikipedia, The Free EncyclopediaDocument3 pagesAcid Phosphatase - Wikipedia, The Free EncyclopediaBinu GeorgeNo ratings yet
- Astaxanthin Manufacturers Cuddalore - Astaxanthin Suppliers Tamil Nadu - AstaxanDocument2 pagesAstaxanthin Manufacturers Cuddalore - Astaxanthin Suppliers Tamil Nadu - AstaxanBinu GeorgeNo ratings yet
- Removal of Dyes From Textile Industry Effluent: A ReviewDocument5 pagesRemoval of Dyes From Textile Industry Effluent: A ReviewBinu GeorgeNo ratings yet
- Molecules: Essential Oils and Their Constituents: Anticonvulsant ActivityDocument17 pagesMolecules: Essential Oils and Their Constituents: Anticonvulsant ActivityBinu GeorgeNo ratings yet
- Hexaconazole - Toxicity, Ecological Toxicity and Regulatory InformationDocument6 pagesHexaconazole - Toxicity, Ecological Toxicity and Regulatory InformationBinu GeorgeNo ratings yet
- 6 Libre PDFDocument1 page6 Libre PDFBinu GeorgeNo ratings yet
- Effect of Endocrine Disruptor Pesticides: A ReviewDocument39 pagesEffect of Endocrine Disruptor Pesticides: A ReviewBinu GeorgeNo ratings yet
- Reproductive Endocrinology FinalDocument98 pagesReproductive Endocrinology FinalChino Paolo SamsonNo ratings yet
- The Concept of Embryonic BreathingDocument43 pagesThe Concept of Embryonic BreathingViktória Molnár100% (3)
- Mid Im 2020 AnatomyDocument178 pagesMid Im 2020 AnatomyXzailheene Dela FuenteNo ratings yet
- Hemorrhage NCPDocument4 pagesHemorrhage NCPElishaNo ratings yet
- Bone TumourDocument78 pagesBone TumourGaluh Kresna BayuNo ratings yet
- Medical Complications of Type 2 DiabetesDocument422 pagesMedical Complications of Type 2 DiabetesMayracpp.16No ratings yet
- Nature Neuroscience February 1999Document94 pagesNature Neuroscience February 1999monomaniahomicidaNo ratings yet
- Legal MedDocument7 pagesLegal MedAlvastro B. LiagaoNo ratings yet
- RiskcalculatorDocument6 pagesRiskcalculatorRenato ValdiviaNo ratings yet
- System 1 Binaural Beats 0001Document12 pagesSystem 1 Binaural Beats 0001Golden SunriseNo ratings yet
- Seminar Format Ece983Document19 pagesSeminar Format Ece983Sheeza MalikNo ratings yet
- Characteristics of Stabilizer Muscles A Systematic ReviewDocument11 pagesCharacteristics of Stabilizer Muscles A Systematic ReviewIvan MuñozNo ratings yet
- Raw Meat vs. Cooked Meat - Which Is HealthierDocument33 pagesRaw Meat vs. Cooked Meat - Which Is HealthierMark Sloan100% (1)
- Chapter 04 Chemical Equilibrium PDFDocument150 pagesChapter 04 Chemical Equilibrium PDFJared Philip Marquez CondezNo ratings yet
- Human Saliva A Potential Diagnostic MediumDocument9 pagesHuman Saliva A Potential Diagnostic MediumInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- AMES TestDocument11 pagesAMES TestJarena Ria ZolinaNo ratings yet
- Biotensegrity ReviewDocument19 pagesBiotensegrity ReviewTensegrity Wiki100% (3)
- The Spiral Engine of Locomotion: Five Primary Kinetic ChainsDocument12 pagesThe Spiral Engine of Locomotion: Five Primary Kinetic ChainsAbhishek VermaNo ratings yet
- Keywords: Q1. Fill in The Blanks: Arteries Red Blood Cells. Capillaries. Heartbeat. Urea. Salts. Urine. Transpiration. Q2. Match UpDocument7 pagesKeywords: Q1. Fill in The Blanks: Arteries Red Blood Cells. Capillaries. Heartbeat. Urea. Salts. Urine. Transpiration. Q2. Match UpGiteshNo ratings yet
- Brain DominanceDocument33 pagesBrain DominanceAndreea Ilie100% (1)
- John Milton Hagen Blood Group SystemDocument5 pagesJohn Milton Hagen Blood Group SystemTia EANo ratings yet
- Physical Assessment Answer 16 PrintedDocument3 pagesPhysical Assessment Answer 16 PrintedJune Dumdumaya100% (2)
- Cells Vocabulary PDFDocument2 pagesCells Vocabulary PDFMoody SaiiNo ratings yet
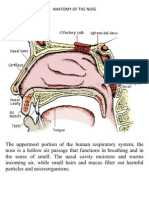
- Anatomy of The NoseDocument2 pagesAnatomy of The NosePablo PasabaNo ratings yet
- 2.1.3.5 - Inkompatibilitas Darah DJDocument14 pages2.1.3.5 - Inkompatibilitas Darah DJnurul ramadhiniNo ratings yet
- Effects of Potassium Levels On Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock SeedlingsDocument13 pagesEffects of Potassium Levels On Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock SeedlingsramdanNo ratings yet
- Introduction and Scope of Anatomy and PhysiologyDocument11 pagesIntroduction and Scope of Anatomy and PhysiologyPrathiNo ratings yet
- Interactive Spinal Cord Atlas: Using The Atlas The GoalDocument13 pagesInteractive Spinal Cord Atlas: Using The Atlas The GoalAffan ElahiNo ratings yet
- Body Buffers and Acid Base BalanceDocument43 pagesBody Buffers and Acid Base BalanceharisNo ratings yet