
Zheng 2006 NatNeuro Bursting Intrinsic Retina PDF
Zheng 2006 NatNeuro Bursting Intrinsic Retina PDF
You might also like
- Osmosis WSDocument2 pagesOsmosis WSSylva Sagita56% (9)
- AUBF Case StudyDocument35 pagesAUBF Case StudyAlecs Elinor Amoguis100% (3)
- Systemic PhysiologyDocument7 pagesSystemic Physiologymichael777No ratings yet
- NROSCI 1012 - Lecture 23Document3 pagesNROSCI 1012 - Lecture 23HonzaNo ratings yet
- CH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewDocument4 pagesCH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewoguzNo ratings yet
- NROSCI 1012 - Lecture 22Document4 pagesNROSCI 1012 - Lecture 22HonzaNo ratings yet
- Brain Res 1977 AghajanianDocument8 pagesBrain Res 1977 Aghajanianndyfk54dpgNo ratings yet
- John Eccles SynapsesDocument22 pagesJohn Eccles SynapsesLeandro XimonNo ratings yet
- KORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked ResponsesDocument5 pagesKORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked Responsesfernando_mart859243No ratings yet
- NROSCI 1012 - Lecture 24Document4 pagesNROSCI 1012 - Lecture 24HonzaNo ratings yet
- Electrical Resonance With Voltage-Gated Ion ChannelsDocument8 pagesElectrical Resonance With Voltage-Gated Ion ChannelsNusrat JahanNo ratings yet
- Myasthenia Gravis - PathophysiologyDocument3 pagesMyasthenia Gravis - PathophysiologyDill BadaróNo ratings yet
- CVS - Cardiac ElectrophysiologyDocument42 pagesCVS - Cardiac Electrophysiologyspitzmark2030No ratings yet
- Characteristics and Plasticity of Electrical SynapDocument16 pagesCharacteristics and Plasticity of Electrical SynapjaneNo ratings yet
- Buzsaki02 Nikolaus ExzerptDocument7 pagesBuzsaki02 Nikolaus ExzerptsiamakNo ratings yet
- Synaptic Depression: Many Different Possible MechanismsDocument24 pagesSynaptic Depression: Many Different Possible MechanismsThe Panda EntertainerNo ratings yet
- Calcium Control of Neurotransmitter Release PDFDocument15 pagesCalcium Control of Neurotransmitter Release PDFNisa Yulianti SuprahmanNo ratings yet
- Calcium Control of Neurotransmitter ReleaseDocument15 pagesCalcium Control of Neurotransmitter ReleaseNisa Yulianti SuprahmanNo ratings yet
- Unit1 BERS Notes-Part2Document11 pagesUnit1 BERS Notes-Part2Mr. PavanNo ratings yet
- Grupo 1 Practica 14Document12 pagesGrupo 1 Practica 14Sadit Chauca VelaNo ratings yet
- 10.3 Muscle Fiber Excitation, Contraction, and Relaxation - Anatomy & PhysiologyDocument1 page10.3 Muscle Fiber Excitation, Contraction, and Relaxation - Anatomy & Physiologyjszkutak24No ratings yet
- The Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesDocument30 pagesThe Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesKhel VincentNo ratings yet
- Börgers - 2017 - An Introduction To Modeling Neuronal Dynamics.Document63 pagesBörgers - 2017 - An Introduction To Modeling Neuronal Dynamics.jorge ignacio moreno fariasNo ratings yet
- Ca 1.3 and BK Channels For Timing and Regulating Cell FiringDocument14 pagesCa 1.3 and BK Channels For Timing and Regulating Cell FiringGabriel L. DebastianiNo ratings yet
- ModelSystem Synapse NeurotransmittersDocument41 pagesModelSystem Synapse NeurotransmittersTapan Kumar PalNo ratings yet
- 1982 Synapses Between L-Neurons J Comp PhysiolDocument14 pages1982 Synapses Between L-Neurons J Comp PhysiolClub OrilyonNo ratings yet
- Cellular Biology of EpileptogenesisDocument10 pagesCellular Biology of EpileptogenesisandreacamachogonNo ratings yet
- Accidental Degeneracy Beats: A Distinct Type of Beat Phenomenon in Nonlinear Optical SpectrosDocument16 pagesAccidental Degeneracy Beats: A Distinct Type of Beat Phenomenon in Nonlinear Optical SpectrosMiguel CervantesNo ratings yet
- Spike Train Analysis and Modelling 1: Christophe PouzatDocument36 pagesSpike Train Analysis and Modelling 1: Christophe PouzatHoyoNo ratings yet
- Skeletal Muscle Physiology PDFDocument6 pagesSkeletal Muscle Physiology PDFAstri Ggamjong Xiao LuNo ratings yet
- Sakai 1988Document19 pagesSakai 1988AnissaNo ratings yet
- Cardio Physiology FullDocument11 pagesCardio Physiology FullSara Lee Wei LiNo ratings yet
- Synapses HistologyDocument6 pagesSynapses HistologySatwant SinghNo ratings yet
- NEUROMUSCULARDocument29 pagesNEUROMUSCULARshikhaNo ratings yet
- Synaptic IntegrationDocument5 pagesSynaptic IntegrationryanNo ratings yet
- Learning Objectives: Chapter 4: Electrophysiology of The HeartDocument28 pagesLearning Objectives: Chapter 4: Electrophysiology of The HeartLaura PucheNo ratings yet
- On The Quantal Mechanism of Neural Transmitter Release: Nobel Lecture, December 12, 1970Document8 pagesOn The Quantal Mechanism of Neural Transmitter Release: Nobel Lecture, December 12, 1970Spk SudhinNo ratings yet
- SynapseDocument45 pagesSynapseroyjason4518No ratings yet
- Advanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Document28 pagesAdvanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Muhammad Mudassar ChudaryNo ratings yet
- Pharmacology of TheDocument21 pagesPharmacology of TheKenneth NuñezNo ratings yet
- Christian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionDocument13 pagesChristian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionFedrmNo ratings yet
- Nervous Coordination WorksheetDocument4 pagesNervous Coordination WorksheetAshley BedassieNo ratings yet
- Principles of Cardiac ElectrophysiologyDocument98 pagesPrinciples of Cardiac ElectrophysiologyAbnet WondimuNo ratings yet
- Jurnal 55Document1 pageJurnal 55rizalmantaNo ratings yet
- CardiacDocument32 pagesCardiacAlli OmerNo ratings yet
- Logy Assighnment On Automaticity Rhythmicity AutorecoveredDocument10 pagesLogy Assighnment On Automaticity Rhythmicity AutorecoveredM Asif RiazNo ratings yet
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Chapter 1Document21 pagesChapter 1Chrisantus OkakaNo ratings yet
- Bahasa Inggris Stahl Ion ChanelDocument5 pagesBahasa Inggris Stahl Ion ChanelMuhammad Zul Fahmi AkbarNo ratings yet
- tmpABFB TMPDocument11 pagestmpABFB TMPFrontiersNo ratings yet
- UNIT 9 NotesDocument5 pagesUNIT 9 NotesMariaNo ratings yet
- Neuromuscular PsysioloyDocument59 pagesNeuromuscular Psysioloypuppy2207100% (2)
- Lab 4Document12 pagesLab 4api-251827644No ratings yet
- Neuromuscular PhysiologyDocument44 pagesNeuromuscular PhysiologyAnkita NigamNo ratings yet
- Key of SEQs Heart, Nerve and Muscle Revision TestDocument11 pagesKey of SEQs Heart, Nerve and Muscle Revision TestMudassar Roomi100% (2)
- Bio 336 Physio Exam 1 EssaysDocument3 pagesBio 336 Physio Exam 1 EssayssheilaNo ratings yet
- Nervous System 2Document4 pagesNervous System 2P KNo ratings yet
- Extrasystoles ParasystolesDocument6 pagesExtrasystoles ParasystolesLivia Maria RaduNo ratings yet
- Cardiac L1CVSDocument19 pagesCardiac L1CVSQutaybah JahmanyNo ratings yet
- Neural Spiking and Synaptic Transmission: Action PotentialDocument6 pagesNeural Spiking and Synaptic Transmission: Action PotentialLoc HuynhNo ratings yet
- Cardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateDocument32 pagesCardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateClaudia AlessandraNo ratings yet
- Science (A Progressive Approach) STDocument9 pagesScience (A Progressive Approach) STAbi ZabalaNo ratings yet
- SPSR 17 P1Document21 pagesSPSR 17 P1Praveen KumarNo ratings yet
- Wonderfallfinal 2Document10 pagesWonderfallfinal 2api-273595156No ratings yet
- Microbes in Human Welfare: SolutionsDocument24 pagesMicrobes in Human Welfare: SolutionsRana GhoshNo ratings yet
- Myasthenia Gravis. SURYADocument41 pagesMyasthenia Gravis. SURYANofilia Citra CandraNo ratings yet
- Examen Grado 2010 Ingles UC3MDocument4 pagesExamen Grado 2010 Ingles UC3MAnonymous mCY1TNHTPyNo ratings yet
- The Implications of Vitamin D Status During Pregnancy On Mother and Her Developing ChildDocument11 pagesThe Implications of Vitamin D Status During Pregnancy On Mother and Her Developing ChildindahNo ratings yet
- Phylum Coelenterata ClassificationDocument3 pagesPhylum Coelenterata ClassificationSudesh Rathod100% (1)
- The World's Flora and FaunaDocument8 pagesThe World's Flora and Faunakleberviteri120No ratings yet
- Achievement Test in Science and Health 6Document5 pagesAchievement Test in Science and Health 6Cess MagtanongNo ratings yet
- The Flowers of Ilex GuayusaDocument7 pagesThe Flowers of Ilex GuayusaGabriela Barriga CoronelNo ratings yet
- Effectiveness of Targetes Erecta Linn As A Natural Serpentes Repellant - EditedDocument3 pagesEffectiveness of Targetes Erecta Linn As A Natural Serpentes Repellant - EditedJameel Jon RifarealNo ratings yet
- The Ecology of Insect Populations Theory and Practice Clark Geier Hughes and MorrisDocument245 pagesThe Ecology of Insect Populations Theory and Practice Clark Geier Hughes and MorrisJulio César Chacón Hernández100% (2)
- SBBMCH 2002Document169 pagesSBBMCH 2002rousesNo ratings yet
- TEMPLATE For Handout in Tecaf 103 1 2 1 1Document5 pagesTEMPLATE For Handout in Tecaf 103 1 2 1 1Nicu CaballeroNo ratings yet
- What Are The Classification of ALP?Document4 pagesWhat Are The Classification of ALP?Frances FranciscoNo ratings yet
- Enzymes Used in DetergentsDocument9 pagesEnzymes Used in DetergentsAlina Baciu100% (3)
- Sakhanokho 2018Document6 pagesSakhanokho 2018AlexNo ratings yet
- Kids Can 3 MaterialDocument9 pagesKids Can 3 MaterialIngrid MuñozNo ratings yet
- ReproductionDocument127 pagesReproductionmohamed salahuddinNo ratings yet
- Facilitating LearningDocument240 pagesFacilitating LearningAthena Cassandra AntonioNo ratings yet
- 11 SCI PC2 BIO T2 2122 - RespirationDocument3 pages11 SCI PC2 BIO T2 2122 - RespirationSharah Nabilla0% (1)
- Om Prakash PPT BiologyDocument11 pagesOm Prakash PPT BiologyOm Prakash100% (1)
- 16732-Article Text-75540-82300-10-20230101Document11 pages16732-Article Text-75540-82300-10-20230101Oksana VedorikaNo ratings yet
- Development of Vertebra, Ribs and Skull 2007Document16 pagesDevelopment of Vertebra, Ribs and Skull 2007Qaiser InayatNo ratings yet
- 1 Kingdom MoneraDocument20 pages1 Kingdom MoneraChrisel Luat Lopez0% (1)
- Molecular Networking As A Drug Discovery, Drug Metabolism, and Precision Medicine StrategyDocument12 pagesMolecular Networking As A Drug Discovery, Drug Metabolism, and Precision Medicine StrategySam SonNo ratings yet
- BiochemistryDocument22 pagesBiochemistryChia JoseNo ratings yet































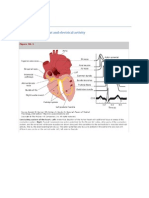














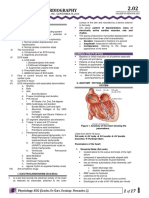






















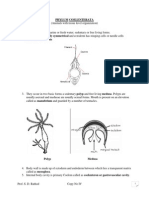




















Download as pdf or txt
You might also like
- Osmosis WSDocument2 pagesOsmosis WSSylva Sagita56% (9)
- AUBF Case StudyDocument35 pagesAUBF Case StudyAlecs Elinor Amoguis100% (3)
- Systemic PhysiologyDocument7 pagesSystemic Physiologymichael777No ratings yet
- NROSCI 1012 - Lecture 23Document3 pagesNROSCI 1012 - Lecture 23HonzaNo ratings yet
- CH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewDocument4 pagesCH5 THE SYNAPSE AND ACTION POTENTIAL GENERATION ON THE POSTSYNAPTIC MEMRANE NewoguzNo ratings yet
- NROSCI 1012 - Lecture 22Document4 pagesNROSCI 1012 - Lecture 22HonzaNo ratings yet
- Brain Res 1977 AghajanianDocument8 pagesBrain Res 1977 Aghajanianndyfk54dpgNo ratings yet
- John Eccles SynapsesDocument22 pagesJohn Eccles SynapsesLeandro XimonNo ratings yet
- KORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked ResponsesDocument5 pagesKORN Et Al - Spontaneous Quantal Currents in A Central Neuron Match Predictions From Binominal Analysis of Evoked Responsesfernando_mart859243No ratings yet
- NROSCI 1012 - Lecture 24Document4 pagesNROSCI 1012 - Lecture 24HonzaNo ratings yet
- Electrical Resonance With Voltage-Gated Ion ChannelsDocument8 pagesElectrical Resonance With Voltage-Gated Ion ChannelsNusrat JahanNo ratings yet
- Myasthenia Gravis - PathophysiologyDocument3 pagesMyasthenia Gravis - PathophysiologyDill BadaróNo ratings yet
- CVS - Cardiac ElectrophysiologyDocument42 pagesCVS - Cardiac Electrophysiologyspitzmark2030No ratings yet
- Characteristics and Plasticity of Electrical SynapDocument16 pagesCharacteristics and Plasticity of Electrical SynapjaneNo ratings yet
- Buzsaki02 Nikolaus ExzerptDocument7 pagesBuzsaki02 Nikolaus ExzerptsiamakNo ratings yet
- Synaptic Depression: Many Different Possible MechanismsDocument24 pagesSynaptic Depression: Many Different Possible MechanismsThe Panda EntertainerNo ratings yet
- Calcium Control of Neurotransmitter Release PDFDocument15 pagesCalcium Control of Neurotransmitter Release PDFNisa Yulianti SuprahmanNo ratings yet
- Calcium Control of Neurotransmitter ReleaseDocument15 pagesCalcium Control of Neurotransmitter ReleaseNisa Yulianti SuprahmanNo ratings yet
- Unit1 BERS Notes-Part2Document11 pagesUnit1 BERS Notes-Part2Mr. PavanNo ratings yet
- Grupo 1 Practica 14Document12 pagesGrupo 1 Practica 14Sadit Chauca VelaNo ratings yet
- 10.3 Muscle Fiber Excitation, Contraction, and Relaxation - Anatomy & PhysiologyDocument1 page10.3 Muscle Fiber Excitation, Contraction, and Relaxation - Anatomy & Physiologyjszkutak24No ratings yet
- The Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesDocument30 pagesThe Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesKhel VincentNo ratings yet
- Börgers - 2017 - An Introduction To Modeling Neuronal Dynamics.Document63 pagesBörgers - 2017 - An Introduction To Modeling Neuronal Dynamics.jorge ignacio moreno fariasNo ratings yet
- Ca 1.3 and BK Channels For Timing and Regulating Cell FiringDocument14 pagesCa 1.3 and BK Channels For Timing and Regulating Cell FiringGabriel L. DebastianiNo ratings yet
- ModelSystem Synapse NeurotransmittersDocument41 pagesModelSystem Synapse NeurotransmittersTapan Kumar PalNo ratings yet
- 1982 Synapses Between L-Neurons J Comp PhysiolDocument14 pages1982 Synapses Between L-Neurons J Comp PhysiolClub OrilyonNo ratings yet
- Cellular Biology of EpileptogenesisDocument10 pagesCellular Biology of EpileptogenesisandreacamachogonNo ratings yet
- Accidental Degeneracy Beats: A Distinct Type of Beat Phenomenon in Nonlinear Optical SpectrosDocument16 pagesAccidental Degeneracy Beats: A Distinct Type of Beat Phenomenon in Nonlinear Optical SpectrosMiguel CervantesNo ratings yet
- Spike Train Analysis and Modelling 1: Christophe PouzatDocument36 pagesSpike Train Analysis and Modelling 1: Christophe PouzatHoyoNo ratings yet
- Skeletal Muscle Physiology PDFDocument6 pagesSkeletal Muscle Physiology PDFAstri Ggamjong Xiao LuNo ratings yet
- Sakai 1988Document19 pagesSakai 1988AnissaNo ratings yet
- Cardio Physiology FullDocument11 pagesCardio Physiology FullSara Lee Wei LiNo ratings yet
- Synapses HistologyDocument6 pagesSynapses HistologySatwant SinghNo ratings yet
- NEUROMUSCULARDocument29 pagesNEUROMUSCULARshikhaNo ratings yet
- Synaptic IntegrationDocument5 pagesSynaptic IntegrationryanNo ratings yet
- Learning Objectives: Chapter 4: Electrophysiology of The HeartDocument28 pagesLearning Objectives: Chapter 4: Electrophysiology of The HeartLaura PucheNo ratings yet
- On The Quantal Mechanism of Neural Transmitter Release: Nobel Lecture, December 12, 1970Document8 pagesOn The Quantal Mechanism of Neural Transmitter Release: Nobel Lecture, December 12, 1970Spk SudhinNo ratings yet
- SynapseDocument45 pagesSynapseroyjason4518No ratings yet
- Advanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Document28 pagesAdvanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Muhammad Mudassar ChudaryNo ratings yet
- Pharmacology of TheDocument21 pagesPharmacology of TheKenneth NuñezNo ratings yet
- Christian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionDocument13 pagesChristian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionFedrmNo ratings yet
- Nervous Coordination WorksheetDocument4 pagesNervous Coordination WorksheetAshley BedassieNo ratings yet
- Principles of Cardiac ElectrophysiologyDocument98 pagesPrinciples of Cardiac ElectrophysiologyAbnet WondimuNo ratings yet
- Jurnal 55Document1 pageJurnal 55rizalmantaNo ratings yet
- CardiacDocument32 pagesCardiacAlli OmerNo ratings yet
- Logy Assighnment On Automaticity Rhythmicity AutorecoveredDocument10 pagesLogy Assighnment On Automaticity Rhythmicity AutorecoveredM Asif RiazNo ratings yet
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Chapter 1Document21 pagesChapter 1Chrisantus OkakaNo ratings yet
- Bahasa Inggris Stahl Ion ChanelDocument5 pagesBahasa Inggris Stahl Ion ChanelMuhammad Zul Fahmi AkbarNo ratings yet
- tmpABFB TMPDocument11 pagestmpABFB TMPFrontiersNo ratings yet
- UNIT 9 NotesDocument5 pagesUNIT 9 NotesMariaNo ratings yet
- Neuromuscular PsysioloyDocument59 pagesNeuromuscular Psysioloypuppy2207100% (2)
- Lab 4Document12 pagesLab 4api-251827644No ratings yet
- Neuromuscular PhysiologyDocument44 pagesNeuromuscular PhysiologyAnkita NigamNo ratings yet
- Key of SEQs Heart, Nerve and Muscle Revision TestDocument11 pagesKey of SEQs Heart, Nerve and Muscle Revision TestMudassar Roomi100% (2)
- Bio 336 Physio Exam 1 EssaysDocument3 pagesBio 336 Physio Exam 1 EssayssheilaNo ratings yet
- Nervous System 2Document4 pagesNervous System 2P KNo ratings yet
- Extrasystoles ParasystolesDocument6 pagesExtrasystoles ParasystolesLivia Maria RaduNo ratings yet
- Cardiac L1CVSDocument19 pagesCardiac L1CVSQutaybah JahmanyNo ratings yet
- Neural Spiking and Synaptic Transmission: Action PotentialDocument6 pagesNeural Spiking and Synaptic Transmission: Action PotentialLoc HuynhNo ratings yet
- Cardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateDocument32 pagesCardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateClaudia AlessandraNo ratings yet
- Science (A Progressive Approach) STDocument9 pagesScience (A Progressive Approach) STAbi ZabalaNo ratings yet
- SPSR 17 P1Document21 pagesSPSR 17 P1Praveen KumarNo ratings yet
- Wonderfallfinal 2Document10 pagesWonderfallfinal 2api-273595156No ratings yet
- Microbes in Human Welfare: SolutionsDocument24 pagesMicrobes in Human Welfare: SolutionsRana GhoshNo ratings yet
- Myasthenia Gravis. SURYADocument41 pagesMyasthenia Gravis. SURYANofilia Citra CandraNo ratings yet
- Examen Grado 2010 Ingles UC3MDocument4 pagesExamen Grado 2010 Ingles UC3MAnonymous mCY1TNHTPyNo ratings yet
- The Implications of Vitamin D Status During Pregnancy On Mother and Her Developing ChildDocument11 pagesThe Implications of Vitamin D Status During Pregnancy On Mother and Her Developing ChildindahNo ratings yet
- Phylum Coelenterata ClassificationDocument3 pagesPhylum Coelenterata ClassificationSudesh Rathod100% (1)
- The World's Flora and FaunaDocument8 pagesThe World's Flora and Faunakleberviteri120No ratings yet
- Achievement Test in Science and Health 6Document5 pagesAchievement Test in Science and Health 6Cess MagtanongNo ratings yet
- The Flowers of Ilex GuayusaDocument7 pagesThe Flowers of Ilex GuayusaGabriela Barriga CoronelNo ratings yet
- Effectiveness of Targetes Erecta Linn As A Natural Serpentes Repellant - EditedDocument3 pagesEffectiveness of Targetes Erecta Linn As A Natural Serpentes Repellant - EditedJameel Jon RifarealNo ratings yet
- The Ecology of Insect Populations Theory and Practice Clark Geier Hughes and MorrisDocument245 pagesThe Ecology of Insect Populations Theory and Practice Clark Geier Hughes and MorrisJulio César Chacón Hernández100% (2)
- SBBMCH 2002Document169 pagesSBBMCH 2002rousesNo ratings yet
- TEMPLATE For Handout in Tecaf 103 1 2 1 1Document5 pagesTEMPLATE For Handout in Tecaf 103 1 2 1 1Nicu CaballeroNo ratings yet
- What Are The Classification of ALP?Document4 pagesWhat Are The Classification of ALP?Frances FranciscoNo ratings yet
- Enzymes Used in DetergentsDocument9 pagesEnzymes Used in DetergentsAlina Baciu100% (3)
- Sakhanokho 2018Document6 pagesSakhanokho 2018AlexNo ratings yet
- Kids Can 3 MaterialDocument9 pagesKids Can 3 MaterialIngrid MuñozNo ratings yet
- ReproductionDocument127 pagesReproductionmohamed salahuddinNo ratings yet
- Facilitating LearningDocument240 pagesFacilitating LearningAthena Cassandra AntonioNo ratings yet
- 11 SCI PC2 BIO T2 2122 - RespirationDocument3 pages11 SCI PC2 BIO T2 2122 - RespirationSharah Nabilla0% (1)
- Om Prakash PPT BiologyDocument11 pagesOm Prakash PPT BiologyOm Prakash100% (1)
- 16732-Article Text-75540-82300-10-20230101Document11 pages16732-Article Text-75540-82300-10-20230101Oksana VedorikaNo ratings yet
- Development of Vertebra, Ribs and Skull 2007Document16 pagesDevelopment of Vertebra, Ribs and Skull 2007Qaiser InayatNo ratings yet
- 1 Kingdom MoneraDocument20 pages1 Kingdom MoneraChrisel Luat Lopez0% (1)
- Molecular Networking As A Drug Discovery, Drug Metabolism, and Precision Medicine StrategyDocument12 pagesMolecular Networking As A Drug Discovery, Drug Metabolism, and Precision Medicine StrategySam SonNo ratings yet
- BiochemistryDocument22 pagesBiochemistryChia JoseNo ratings yet