
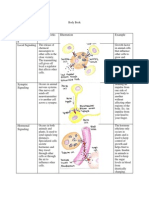
Epigenetic A
Epigenetic A
You might also like
- CH 18 Reading Guide - AnswersDocument6 pagesCH 18 Reading Guide - AnswersSylvia Grace0% (12)
- Acetate-Mediated Novel Survival Strategy Against Drought in Plants-3Document7 pagesAcetate-Mediated Novel Survival Strategy Against Drought in Plants-3MdMezanurRahmanShaonNo ratings yet
- Chapter 19 LodishDocument52 pagesChapter 19 LodishablucNo ratings yet
- Globin Gene & Molecular Clock PDFDocument36 pagesGlobin Gene & Molecular Clock PDFManisha BishtNo ratings yet
- Development Q&ADocument6 pagesDevelopment Q&AKathleen AraújoNo ratings yet
- bbt193003 - Yeshua ShahzadDocument9 pagesbbt193003 - Yeshua ShahzadAbuzar KhanNo ratings yet
- Differential Gene Expression and DevelopmentDocument9 pagesDifferential Gene Expression and DevelopmentShaher Bano Mirza100% (1)
- 10.4: Eukaryotic Gene RegulationDocument7 pages10.4: Eukaryotic Gene RegulationFlorence Gaone GaongalelweNo ratings yet
- HowgenesworkDocument20 pagesHowgenesworkklundermadona93No ratings yet
- How Genes WorkDocument20 pagesHow Genes WorkSunnyNo ratings yet
- Epigenetics and Well-Being: Optimal Adaptation To The EnvironmentDocument21 pagesEpigenetics and Well-Being: Optimal Adaptation To The EnvironmentOlivier BenarrocheNo ratings yet
- Gene ExpressionDocument16 pagesGene ExpressionmidhunbonumahanthiNo ratings yet
- 7 Gendev PDFDocument12 pages7 Gendev PDFJohn DeHansNo ratings yet
- Gene Protein and RegulationDocument4 pagesGene Protein and Regulationapi-248290141No ratings yet
- Cell Communicatio N How It Works Illustration ExampleDocument8 pagesCell Communicatio N How It Works Illustration ExampleMaha HusseiniNo ratings yet
- Signalling in CellsDocument11 pagesSignalling in CellsAditi JhaNo ratings yet
- Gene Regulation - Operon Theory - Microbiology - OpenStaxDocument12 pagesGene Regulation - Operon Theory - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- Network Motifs: Theory and Experimental Approaches: ReviewsDocument12 pagesNetwork Motifs: Theory and Experimental Approaches: ReviewsIndra Prakash JhaNo ratings yet
- Chapter 11 Gene ExpressionDocument17 pagesChapter 11 Gene ExpressionAllyssa Mae ClorionNo ratings yet
- Name: Ricki Elaine Dalipe Sched: Wed 1-6pmDocument4 pagesName: Ricki Elaine Dalipe Sched: Wed 1-6pmZeid ZiedNo ratings yet
- Self-Organization and The Origins of Life: The Managed-Metabolism HypothesisDocument24 pagesSelf-Organization and The Origins of Life: The Managed-Metabolism HypothesisEnaNo ratings yet
- Notes 221210 003035Document3 pagesNotes 221210 003035SWAGGANG 15No ratings yet
- Questions and Answers For Scientific Article June 2011Document14 pagesQuestions and Answers For Scientific Article June 2011Zydhan ZahidNo ratings yet
- Cell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicDocument6 pagesCell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicMohan RaajNo ratings yet
- GENETIKADocument19 pagesGENETIKANur AfifahizaNo ratings yet
- Epigenetics Book: The Most Comprehensive Exploration of the Practical, Social and Ethical Impact of DNA on Our Society and Our WorldFrom EverandEpigenetics Book: The Most Comprehensive Exploration of the Practical, Social and Ethical Impact of DNA on Our Society and Our WorldNo ratings yet
- Wolpert, L. - 1994 - Do We Understand DevelopmentDocument2 pagesWolpert, L. - 1994 - Do We Understand DevelopmentcontulmmivNo ratings yet
- Unit 2 QuestionsDocument3 pagesUnit 2 QuestionssseettNo ratings yet
- Handouts Gen. Bio Oct 3 6 2022Document14 pagesHandouts Gen. Bio Oct 3 6 2022Mark Kenneth CastilloNo ratings yet
- Ch18 Gene Expression PPT NotesDocument29 pagesCh18 Gene Expression PPT NotesChorusa QuinnNo ratings yet
- Module 6 - Genetics, Evolution and EcosystemsDocument65 pagesModule 6 - Genetics, Evolution and Ecosystemsanonuser854No ratings yet
- All Sexual Organisms Begin As A Single CellDocument12 pagesAll Sexual Organisms Begin As A Single CellAFRIADMA AULIA PERDANANo ratings yet
- Genetics (Introduction)Document10 pagesGenetics (Introduction)Mielah RuthNo ratings yet
- Teacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyDocument17 pagesTeacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyApratim SinghNo ratings yet
- ARTICULO 1cell Signallingcornell2005Document4 pagesARTICULO 1cell Signallingcornell2005Carmen MaRiNo ratings yet
- pq002776 PDFDocument3 pagespq002776 PDFDavid Espinosa SuárezNo ratings yet
- Insights New111Document13 pagesInsights New111cassidy conchaNo ratings yet
- Proteins Can Be Classified Into Two Broad TypesDocument19 pagesProteins Can Be Classified Into Two Broad TypesHelmi ZztNo ratings yet
- Chapter 16 - Gene ExpressionDocument20 pagesChapter 16 - Gene ExpressionAyushi MauryaNo ratings yet
- Running Head: Stem Cells Biology 1Document5 pagesRunning Head: Stem Cells Biology 1Donslay Donnell DonNo ratings yet
- Cell Cycle: Synthesis Phase and Second GapDocument4 pagesCell Cycle: Synthesis Phase and Second GapJhude JosephNo ratings yet
- Generic Master ChefsDocument11 pagesGeneric Master ChefsAquib AmirNo ratings yet
- Unit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionDocument76 pagesUnit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionMuhammad SibtainNo ratings yet
- Chromatin Modi Cations and Their FunctionDocument13 pagesChromatin Modi Cations and Their FunctionualigolNo ratings yet
- Chapter 1: General Principles of Cellular OrganizationDocument9 pagesChapter 1: General Principles of Cellular OrganizationWissam MoslehNo ratings yet
- Cell Biology: SubscribeDocument2 pagesCell Biology: SubscribeAnonymous fKViHeaw100% (1)
- TransposonsDocument20 pagesTransposonsprerna tripathyNo ratings yet
- Cell Cyle Discussion QuestionsDocument4 pagesCell Cyle Discussion QuestionsHamed NilforoshanNo ratings yet
- Answers by Stafford Valentine Redden, VIHS, Head of Biology 1Document14 pagesAnswers by Stafford Valentine Redden, VIHS, Head of Biology 1areyouthere92No ratings yet
- Please Explain What Is Gene Expression (Please Add Diagram and Example, If Possible)Document2 pagesPlease Explain What Is Gene Expression (Please Add Diagram and Example, If Possible)devaNo ratings yet
- Signaling Molecules and Their ReceptorsDocument13 pagesSignaling Molecules and Their ReceptorsShweta SinghNo ratings yet
- Cell Differentiation: Main PrinciplesDocument44 pagesCell Differentiation: Main PrinciplesZainab Jamal SiddiquiNo ratings yet
- Developmental GeneticsDocument20 pagesDevelopmental Geneticsabhishek100% (1)
- Informe EpigeneticaDocument22 pagesInforme EpigeneticaHanyo plpNo ratings yet
- Meiosis PDFDocument54 pagesMeiosis PDFYusrahanna daulayNo ratings yet
- Gene ExpressionDocument29 pagesGene ExpressionChorusa QuinnNo ratings yet
- A Third Way: More Articles On EvolutionDocument5 pagesA Third Way: More Articles On EvolutionjlventiganNo ratings yet
- Guangjin Pan Et Al - A Negative Feedback Loop of Transcription Factors That Controls Stem Cell Pluripotency and Self-RenewalDocument12 pagesGuangjin Pan Et Al - A Negative Feedback Loop of Transcription Factors That Controls Stem Cell Pluripotency and Self-RenewalGmewop30mNo ratings yet
- Genetic ExpressionsDocument6 pagesGenetic ExpressionsRobertaNo ratings yet
- Role of Cytoplasm in Cell DifferentiationDocument7 pagesRole of Cytoplasm in Cell DifferentiationJ LALNo ratings yet
- Beyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchFrom EverandBeyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchNo ratings yet
- 03 Genetics A3 Revision-Sheet A3formatms - Pdfviewer.action DownloadDocument1 page03 Genetics A3 Revision-Sheet A3formatms - Pdfviewer.action DownloadSanaa SamkoNo ratings yet
- Biochemical Energy ProductionDocument5 pagesBiochemical Energy ProductionAlexandria StylesNo ratings yet
- The Metabolism Booster Complex Helps Optimise Cellular Function and Glucose RegulationDocument2 pagesThe Metabolism Booster Complex Helps Optimise Cellular Function and Glucose RegulationCecile SiauwNo ratings yet
- Lippincotts Illustrated Biochem Review 4th Ed Compiled QuestionsDocument36 pagesLippincotts Illustrated Biochem Review 4th Ed Compiled QuestionsKathleen Tubera0% (1)
- RNA Structure, Function, and Synthesis RNADocument6 pagesRNA Structure, Function, and Synthesis RNAMohini BajajNo ratings yet
- EnzymesDocument21 pagesEnzymesCatherine Merilleno100% (1)
- Liver and CurcuminDocument6 pagesLiver and CurcuminLouie Mae SantosNo ratings yet
- DNA FingerprintingDocument20 pagesDNA FingerprintingCleo Poulos100% (1)
- Barba Et Al-2014-Molecular Ecology ResourcesDocument18 pagesBarba Et Al-2014-Molecular Ecology ResourcesAndrés FrankowNo ratings yet
- Computational Approach in Drug Discovery Molecular Docking Using Hex 4.5RDocument7 pagesComputational Approach in Drug Discovery Molecular Docking Using Hex 4.5RSharad JaiswalNo ratings yet
- Genetic Engineering PDFDocument50 pagesGenetic Engineering PDFBrian Rey L. AbingNo ratings yet
- Genetics A Conceptual Approach 5th Edition Pierce Solutions ManualDocument25 pagesGenetics A Conceptual Approach 5th Edition Pierce Solutions ManualJackCoxmozj100% (50)
- L-Glutamine and Nutritional BenefitsDocument8 pagesL-Glutamine and Nutritional BenefitsKevin KiplangatNo ratings yet
- NopigmerinDocument21 pagesNopigmerinInstituto Ricco PortoNo ratings yet
- 2017 Anti-Wrinkle - and - Skin - Turnover - Improvement - EffectsDocument12 pages2017 Anti-Wrinkle - and - Skin - Turnover - Improvement - EffectsVita BūdvytėNo ratings yet
- Chapter 5 The Fundamental Unit of LifeDocument18 pagesChapter 5 The Fundamental Unit of LifeTownship TownshipNo ratings yet
- A Level Biology Essay Model AnswersDocument4 pagesA Level Biology Essay Model AnswersKFCStadivari100% (1)
- EN EpiTect MethyLight PCR HandbookDocument36 pagesEN EpiTect MethyLight PCR HandbookRiska Dwy NurainyNo ratings yet
- Worksheet Ap01Document11 pagesWorksheet Ap01Abhishek SuranaNo ratings yet
- Biosynthesis and Pharmacognosy Lecture 2 - Biosynthesis of CarbohydratesDocument19 pagesBiosynthesis and Pharmacognosy Lecture 2 - Biosynthesis of CarbohydratesFrogWarriorNo ratings yet
- Pharmacology AntidepresantDocument17 pagesPharmacology Antidepresantandhita96No ratings yet
- Nonalcoholic Steatohepatitis DR Harbans SinghDocument3 pagesNonalcoholic Steatohepatitis DR Harbans SinghHarbans Singh100% (1)
- MY 1st Sem. Presttn DNA Damage & Repair PPT MW 2003Document26 pagesMY 1st Sem. Presttn DNA Damage & Repair PPT MW 2003Samhita Kalita100% (1)
- B.pharma 2 SemDocument13 pagesB.pharma 2 Semabhishek.thokeNo ratings yet
- IB DP Biology 1.4 Membrane Transport QuestionDocument19 pagesIB DP Biology 1.4 Membrane Transport QuestionikaNo ratings yet
- Phytocannabinoids:: What Is On The Horizon?Document16 pagesPhytocannabinoids:: What Is On The Horizon?detaciNo ratings yet
- Paracetamol-Induced Renal Tubular Injury: A Role For ER StressDocument10 pagesParacetamol-Induced Renal Tubular Injury: A Role For ER StressمحمدأميندماجNo ratings yet
- L - 80148 LDL.01Document1 pageL - 80148 LDL.01mrashrafiNo ratings yet
























































































Download as pdf or txt
You might also like
- CH 18 Reading Guide - AnswersDocument6 pagesCH 18 Reading Guide - AnswersSylvia Grace0% (12)
- Acetate-Mediated Novel Survival Strategy Against Drought in Plants-3Document7 pagesAcetate-Mediated Novel Survival Strategy Against Drought in Plants-3MdMezanurRahmanShaonNo ratings yet
- Chapter 19 LodishDocument52 pagesChapter 19 LodishablucNo ratings yet
- Globin Gene & Molecular Clock PDFDocument36 pagesGlobin Gene & Molecular Clock PDFManisha BishtNo ratings yet
- Development Q&ADocument6 pagesDevelopment Q&AKathleen AraújoNo ratings yet
- bbt193003 - Yeshua ShahzadDocument9 pagesbbt193003 - Yeshua ShahzadAbuzar KhanNo ratings yet
- Differential Gene Expression and DevelopmentDocument9 pagesDifferential Gene Expression and DevelopmentShaher Bano Mirza100% (1)
- 10.4: Eukaryotic Gene RegulationDocument7 pages10.4: Eukaryotic Gene RegulationFlorence Gaone GaongalelweNo ratings yet
- HowgenesworkDocument20 pagesHowgenesworkklundermadona93No ratings yet
- How Genes WorkDocument20 pagesHow Genes WorkSunnyNo ratings yet
- Epigenetics and Well-Being: Optimal Adaptation To The EnvironmentDocument21 pagesEpigenetics and Well-Being: Optimal Adaptation To The EnvironmentOlivier BenarrocheNo ratings yet
- Gene ExpressionDocument16 pagesGene ExpressionmidhunbonumahanthiNo ratings yet
- 7 Gendev PDFDocument12 pages7 Gendev PDFJohn DeHansNo ratings yet
- Gene Protein and RegulationDocument4 pagesGene Protein and Regulationapi-248290141No ratings yet
- Cell Communicatio N How It Works Illustration ExampleDocument8 pagesCell Communicatio N How It Works Illustration ExampleMaha HusseiniNo ratings yet
- Signalling in CellsDocument11 pagesSignalling in CellsAditi JhaNo ratings yet
- Gene Regulation - Operon Theory - Microbiology - OpenStaxDocument12 pagesGene Regulation - Operon Theory - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- Network Motifs: Theory and Experimental Approaches: ReviewsDocument12 pagesNetwork Motifs: Theory and Experimental Approaches: ReviewsIndra Prakash JhaNo ratings yet
- Chapter 11 Gene ExpressionDocument17 pagesChapter 11 Gene ExpressionAllyssa Mae ClorionNo ratings yet
- Name: Ricki Elaine Dalipe Sched: Wed 1-6pmDocument4 pagesName: Ricki Elaine Dalipe Sched: Wed 1-6pmZeid ZiedNo ratings yet
- Self-Organization and The Origins of Life: The Managed-Metabolism HypothesisDocument24 pagesSelf-Organization and The Origins of Life: The Managed-Metabolism HypothesisEnaNo ratings yet
- Notes 221210 003035Document3 pagesNotes 221210 003035SWAGGANG 15No ratings yet
- Questions and Answers For Scientific Article June 2011Document14 pagesQuestions and Answers For Scientific Article June 2011Zydhan ZahidNo ratings yet
- Cell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicDocument6 pagesCell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicMohan RaajNo ratings yet
- GENETIKADocument19 pagesGENETIKANur AfifahizaNo ratings yet
- Epigenetics Book: The Most Comprehensive Exploration of the Practical, Social and Ethical Impact of DNA on Our Society and Our WorldFrom EverandEpigenetics Book: The Most Comprehensive Exploration of the Practical, Social and Ethical Impact of DNA on Our Society and Our WorldNo ratings yet
- Wolpert, L. - 1994 - Do We Understand DevelopmentDocument2 pagesWolpert, L. - 1994 - Do We Understand DevelopmentcontulmmivNo ratings yet
- Unit 2 QuestionsDocument3 pagesUnit 2 QuestionssseettNo ratings yet
- Handouts Gen. Bio Oct 3 6 2022Document14 pagesHandouts Gen. Bio Oct 3 6 2022Mark Kenneth CastilloNo ratings yet
- Ch18 Gene Expression PPT NotesDocument29 pagesCh18 Gene Expression PPT NotesChorusa QuinnNo ratings yet
- Module 6 - Genetics, Evolution and EcosystemsDocument65 pagesModule 6 - Genetics, Evolution and Ecosystemsanonuser854No ratings yet
- All Sexual Organisms Begin As A Single CellDocument12 pagesAll Sexual Organisms Begin As A Single CellAFRIADMA AULIA PERDANANo ratings yet
- Genetics (Introduction)Document10 pagesGenetics (Introduction)Mielah RuthNo ratings yet
- Teacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyDocument17 pagesTeacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyApratim SinghNo ratings yet
- ARTICULO 1cell Signallingcornell2005Document4 pagesARTICULO 1cell Signallingcornell2005Carmen MaRiNo ratings yet
- pq002776 PDFDocument3 pagespq002776 PDFDavid Espinosa SuárezNo ratings yet
- Insights New111Document13 pagesInsights New111cassidy conchaNo ratings yet
- Proteins Can Be Classified Into Two Broad TypesDocument19 pagesProteins Can Be Classified Into Two Broad TypesHelmi ZztNo ratings yet
- Chapter 16 - Gene ExpressionDocument20 pagesChapter 16 - Gene ExpressionAyushi MauryaNo ratings yet
- Running Head: Stem Cells Biology 1Document5 pagesRunning Head: Stem Cells Biology 1Donslay Donnell DonNo ratings yet
- Cell Cycle: Synthesis Phase and Second GapDocument4 pagesCell Cycle: Synthesis Phase and Second GapJhude JosephNo ratings yet
- Generic Master ChefsDocument11 pagesGeneric Master ChefsAquib AmirNo ratings yet
- Unit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionDocument76 pagesUnit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionMuhammad SibtainNo ratings yet
- Chromatin Modi Cations and Their FunctionDocument13 pagesChromatin Modi Cations and Their FunctionualigolNo ratings yet
- Chapter 1: General Principles of Cellular OrganizationDocument9 pagesChapter 1: General Principles of Cellular OrganizationWissam MoslehNo ratings yet
- Cell Biology: SubscribeDocument2 pagesCell Biology: SubscribeAnonymous fKViHeaw100% (1)
- TransposonsDocument20 pagesTransposonsprerna tripathyNo ratings yet
- Cell Cyle Discussion QuestionsDocument4 pagesCell Cyle Discussion QuestionsHamed NilforoshanNo ratings yet
- Answers by Stafford Valentine Redden, VIHS, Head of Biology 1Document14 pagesAnswers by Stafford Valentine Redden, VIHS, Head of Biology 1areyouthere92No ratings yet
- Please Explain What Is Gene Expression (Please Add Diagram and Example, If Possible)Document2 pagesPlease Explain What Is Gene Expression (Please Add Diagram and Example, If Possible)devaNo ratings yet
- Signaling Molecules and Their ReceptorsDocument13 pagesSignaling Molecules and Their ReceptorsShweta SinghNo ratings yet
- Cell Differentiation: Main PrinciplesDocument44 pagesCell Differentiation: Main PrinciplesZainab Jamal SiddiquiNo ratings yet
- Developmental GeneticsDocument20 pagesDevelopmental Geneticsabhishek100% (1)
- Informe EpigeneticaDocument22 pagesInforme EpigeneticaHanyo plpNo ratings yet
- Meiosis PDFDocument54 pagesMeiosis PDFYusrahanna daulayNo ratings yet
- Gene ExpressionDocument29 pagesGene ExpressionChorusa QuinnNo ratings yet
- A Third Way: More Articles On EvolutionDocument5 pagesA Third Way: More Articles On EvolutionjlventiganNo ratings yet
- Guangjin Pan Et Al - A Negative Feedback Loop of Transcription Factors That Controls Stem Cell Pluripotency and Self-RenewalDocument12 pagesGuangjin Pan Et Al - A Negative Feedback Loop of Transcription Factors That Controls Stem Cell Pluripotency and Self-RenewalGmewop30mNo ratings yet
- Genetic ExpressionsDocument6 pagesGenetic ExpressionsRobertaNo ratings yet
- Role of Cytoplasm in Cell DifferentiationDocument7 pagesRole of Cytoplasm in Cell DifferentiationJ LALNo ratings yet
- Beyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchFrom EverandBeyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchNo ratings yet
- 03 Genetics A3 Revision-Sheet A3formatms - Pdfviewer.action DownloadDocument1 page03 Genetics A3 Revision-Sheet A3formatms - Pdfviewer.action DownloadSanaa SamkoNo ratings yet
- Biochemical Energy ProductionDocument5 pagesBiochemical Energy ProductionAlexandria StylesNo ratings yet
- The Metabolism Booster Complex Helps Optimise Cellular Function and Glucose RegulationDocument2 pagesThe Metabolism Booster Complex Helps Optimise Cellular Function and Glucose RegulationCecile SiauwNo ratings yet
- Lippincotts Illustrated Biochem Review 4th Ed Compiled QuestionsDocument36 pagesLippincotts Illustrated Biochem Review 4th Ed Compiled QuestionsKathleen Tubera0% (1)
- RNA Structure, Function, and Synthesis RNADocument6 pagesRNA Structure, Function, and Synthesis RNAMohini BajajNo ratings yet
- EnzymesDocument21 pagesEnzymesCatherine Merilleno100% (1)
- Liver and CurcuminDocument6 pagesLiver and CurcuminLouie Mae SantosNo ratings yet
- DNA FingerprintingDocument20 pagesDNA FingerprintingCleo Poulos100% (1)
- Barba Et Al-2014-Molecular Ecology ResourcesDocument18 pagesBarba Et Al-2014-Molecular Ecology ResourcesAndrés FrankowNo ratings yet
- Computational Approach in Drug Discovery Molecular Docking Using Hex 4.5RDocument7 pagesComputational Approach in Drug Discovery Molecular Docking Using Hex 4.5RSharad JaiswalNo ratings yet
- Genetic Engineering PDFDocument50 pagesGenetic Engineering PDFBrian Rey L. AbingNo ratings yet
- Genetics A Conceptual Approach 5th Edition Pierce Solutions ManualDocument25 pagesGenetics A Conceptual Approach 5th Edition Pierce Solutions ManualJackCoxmozj100% (50)
- L-Glutamine and Nutritional BenefitsDocument8 pagesL-Glutamine and Nutritional BenefitsKevin KiplangatNo ratings yet
- NopigmerinDocument21 pagesNopigmerinInstituto Ricco PortoNo ratings yet
- 2017 Anti-Wrinkle - and - Skin - Turnover - Improvement - EffectsDocument12 pages2017 Anti-Wrinkle - and - Skin - Turnover - Improvement - EffectsVita BūdvytėNo ratings yet
- Chapter 5 The Fundamental Unit of LifeDocument18 pagesChapter 5 The Fundamental Unit of LifeTownship TownshipNo ratings yet
- A Level Biology Essay Model AnswersDocument4 pagesA Level Biology Essay Model AnswersKFCStadivari100% (1)
- EN EpiTect MethyLight PCR HandbookDocument36 pagesEN EpiTect MethyLight PCR HandbookRiska Dwy NurainyNo ratings yet
- Worksheet Ap01Document11 pagesWorksheet Ap01Abhishek SuranaNo ratings yet
- Biosynthesis and Pharmacognosy Lecture 2 - Biosynthesis of CarbohydratesDocument19 pagesBiosynthesis and Pharmacognosy Lecture 2 - Biosynthesis of CarbohydratesFrogWarriorNo ratings yet
- Pharmacology AntidepresantDocument17 pagesPharmacology Antidepresantandhita96No ratings yet
- Nonalcoholic Steatohepatitis DR Harbans SinghDocument3 pagesNonalcoholic Steatohepatitis DR Harbans SinghHarbans Singh100% (1)
- MY 1st Sem. Presttn DNA Damage & Repair PPT MW 2003Document26 pagesMY 1st Sem. Presttn DNA Damage & Repair PPT MW 2003Samhita Kalita100% (1)
- B.pharma 2 SemDocument13 pagesB.pharma 2 Semabhishek.thokeNo ratings yet
- IB DP Biology 1.4 Membrane Transport QuestionDocument19 pagesIB DP Biology 1.4 Membrane Transport QuestionikaNo ratings yet
- Phytocannabinoids:: What Is On The Horizon?Document16 pagesPhytocannabinoids:: What Is On The Horizon?detaciNo ratings yet
- Paracetamol-Induced Renal Tubular Injury: A Role For ER StressDocument10 pagesParacetamol-Induced Renal Tubular Injury: A Role For ER StressمحمدأميندماجNo ratings yet
- L - 80148 LDL.01Document1 pageL - 80148 LDL.01mrashrafiNo ratings yet