
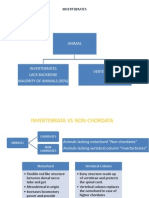
Sponges
Sponges
You might also like
- Fundamentals of Invertebrate Palaeontology MacrofossilsDocument411 pagesFundamentals of Invertebrate Palaeontology MacrofossilsLuthfan100% (2)
- BiologyDocument140 pagesBiologyZandile MuwaloNo ratings yet
- Chapter 4 - Tissues Glands and MembranesDocument49 pagesChapter 4 - Tissues Glands and MembranesNess G. Mamasabulod100% (1)
- Sponges and PlacozoansDocument30 pagesSponges and Placozoansdayahfarday100% (1)
- The Arthropods: Blueprint For SuccessDocument68 pagesThe Arthropods: Blueprint For SuccessKlym CaballasNo ratings yet
- Phylum Porifera, Characteristics and Class ExamplesDocument4 pagesPhylum Porifera, Characteristics and Class ExamplesRobert BarrettoNo ratings yet
- Phylum PoriferaDocument18 pagesPhylum Poriferagonoles81No ratings yet
- Chapter6-Phylum Porifera NotesDocument4 pagesChapter6-Phylum Porifera Notesapi-195601294No ratings yet
- Classification of ProtozoaDocument1 pageClassification of ProtozoaSalil ShauNo ratings yet
- Biology PDFDocument344 pagesBiology PDFUrmilaAnantNo ratings yet
- Tissues NotesDocument7 pagesTissues Noteskapil100% (1)
- Lab Practical 1Document21 pagesLab Practical 1randy_sanchez_10No ratings yet
- 11.2 Domain Bacteria and ArchaeaDocument39 pages11.2 Domain Bacteria and ArchaeaZkdlin SpaceNo ratings yet
- Plant and Animal CellDocument7 pagesPlant and Animal CellHera Lopez100% (1)
- Frog Lab 1Document4 pagesFrog Lab 1api-296981910No ratings yet
- PoriferaDocument39 pagesPoriferaabhishek.ajmaniNo ratings yet
- Porifera - EvolucionDocument9 pagesPorifera - EvolucionJuan David HPNo ratings yet
- Histo Trans Lec 3 1 PDFDocument8 pagesHisto Trans Lec 3 1 PDFBreville SageNo ratings yet
- Types of BlastulaDocument3 pagesTypes of BlastulaDeepanshu Saini80% (5)
- Lecture - Microbial Diversity The Eukaryotic Microbes - Chapter 4 - BSEDDocument5 pagesLecture - Microbial Diversity The Eukaryotic Microbes - Chapter 4 - BSEDMaden betoNo ratings yet
- Exercise 1 The Cell 1Document6 pagesExercise 1 The Cell 1MJMadlangbayan100% (1)
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Phylum Porifera NotesDocument2 pagesPhylum Porifera Notesapi-24708413675% (4)
- A. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItDocument9 pagesA. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItFrancis100% (1)
- Echinostoma IlocanumDocument3 pagesEchinostoma IlocanumChristie Raye NapuliNo ratings yet
- Phylum EchiuraDocument22 pagesPhylum EchiuraEthyl50% (2)
- Homeostasis and The Excretory SystemDocument21 pagesHomeostasis and The Excretory Systemmark smithNo ratings yet
- Stem Cells in HumansDocument4 pagesStem Cells in HumansSana NainaNo ratings yet
- General Classification (All Phylum)Document16 pagesGeneral Classification (All Phylum)kiran PoudelNo ratings yet
- Phylum Nematoda 13Document132 pagesPhylum Nematoda 13Iseth ISeth100% (1)
- CHAPTER 4 BOTANY - Review Questions and AnswersDocument7 pagesCHAPTER 4 BOTANY - Review Questions and AnswersPam GarciaNo ratings yet
- AnimaldistributioniiiDocument22 pagesAnimaldistributioniiiManoo BiliNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- 11.5 Domain Eukarya: Kingdom PlantaeDocument34 pages11.5 Domain Eukarya: Kingdom PlantaeMuhammad HusniNo ratings yet
- ProtozoansDocument2 pagesProtozoansJonathanPayongayongNo ratings yet
- Animal Nutrition: AP BiologyDocument34 pagesAnimal Nutrition: AP BiologyDoods Galdo100% (1)
- CBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsDocument10 pagesCBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsPriyanshu BhadanaNo ratings yet
- Intestinal and Commensal AmoebaDocument9 pagesIntestinal and Commensal AmoebaFuture TrekingNo ratings yet
- Venn Diagram On Osmosis and DiffusionDocument1 pageVenn Diagram On Osmosis and DiffusioncolacastellNo ratings yet
- Plant CellDocument3 pagesPlant CellSUZAIMAHNo ratings yet
- AMOEBADocument74 pagesAMOEBAhuyenthanh1807No ratings yet
- Cell Structure Function and PropertiesDocument10 pagesCell Structure Function and PropertiesWanda JohnNo ratings yet
- The Primary and Secondary Growth of PlantDocument31 pagesThe Primary and Secondary Growth of PlantNOFAIZAH RAMLINo ratings yet
- Animal Kingdom: Basis of ClassificationDocument15 pagesAnimal Kingdom: Basis of ClassificationChaitali Deshmukh100% (1)
- Cell - Histology Trans Part 1&2Document6 pagesCell - Histology Trans Part 1&2Mark AbrazaldoNo ratings yet
- The Species Concept-1Document5 pagesThe Species Concept-1O me khanNo ratings yet
- Activity 9 Phylum NematodaDocument18 pagesActivity 9 Phylum Nematodanicole syNo ratings yet
- Endocrine SystemDocument27 pagesEndocrine Systemcardos cherry100% (1)
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Histology of Gastrointestinal TractDocument46 pagesHistology of Gastrointestinal Tractandreas kevinNo ratings yet
- Animal Tissues 1Document45 pagesAnimal Tissues 1sirajNo ratings yet
- S.3 Biology Vol. 4 Excretion and HomeostasisDocument13 pagesS.3 Biology Vol. 4 Excretion and HomeostasisFrancheli BautistaNo ratings yet
- Plant KingdomDocument16 pagesPlant Kingdomshriyansh singhaniaNo ratings yet
- 3 ClearingDocument3 pages3 ClearingAngel RamosNo ratings yet
- Leukocytes, Granulocytes, The Monocyte-Macrophage System, and InflammationDocument10 pagesLeukocytes, Granulocytes, The Monocyte-Macrophage System, and InflammationShi no MeNo ratings yet
- Chapter 9 The Immune System NotesDocument8 pagesChapter 9 The Immune System NotesFelix LeNo ratings yet
- 55 16saczo1 2020121910192848Document30 pages55 16saczo1 2020121910192848Dandy H HerkoNo ratings yet
- BSC Part I Canal System in PoriferaDocument6 pagesBSC Part I Canal System in PoriferaSriram MNo ratings yet
- 157 18ZC101 2020120608325782Document18 pages157 18ZC101 2020120608325782pm102382No ratings yet
- PoriferaDocument33 pagesPoriferaRushikesh pawar100% (1)
- Porifera ReviewerDocument8 pagesPorifera ReviewerCarlo MendozaNo ratings yet
- Practical Guide Invertebrates PDFDocument61 pagesPractical Guide Invertebrates PDFMOUSTAFA ALEMAMNo ratings yet
- Guia Visual de Organismos de Aguas Profundas Del Caribe Colombiano / Visual Guide of Deep Sea Organisms of The Colombian CaribbeanDocument99 pagesGuia Visual de Organismos de Aguas Profundas Del Caribe Colombiano / Visual Guide of Deep Sea Organisms of The Colombian CaribbeanAlejandro GarciaNo ratings yet
- Honesty Is The Best Policy EssayDocument8 pagesHonesty Is The Best Policy Essaysmgcjvwhd100% (2)
- Feart 10 914319Document12 pagesFeart 10 914319Satrio Hani SamudraNo ratings yet
- SpongesDocument16 pagesSpongesKazuki Fuchoin100% (1)
- PoriferaDocument33 pagesPoriferaRushikesh pawar100% (1)
- 1st August (Porifera)Document7 pages1st August (Porifera)tayyabuetlhrNo ratings yet
- Chapter 9 - ZoologyDocument3 pagesChapter 9 - ZoologyAngel MerilloNo ratings yet
- Phylum Cnidaria and Porifera Group 1Document32 pagesPhylum Cnidaria and Porifera Group 1JulesNo ratings yet
- PoriferaDocument11 pagesPoriferaJohn AdamNo ratings yet
- Sponge ColoringDocument2 pagesSponge ColoringMathias CronqvistNo ratings yet
- Practical Manual in Zoology: Dr. A. AmsathDocument323 pagesPractical Manual in Zoology: Dr. A. AmsathAnnapoorna SagarNo ratings yet
- The Fauna Associated With Outer Shelf and Upper Slope SpongesDocument22 pagesThe Fauna Associated With Outer Shelf and Upper Slope SpongesPedro Henrique ClerierNo ratings yet
- Porififera and CnidariaDocument6 pagesPorififera and CnidariaFlorence PalomarNo ratings yet
- Module 8 - Multi-Cellular and Tissue Levels of OrganizationDocument25 pagesModule 8 - Multi-Cellular and Tissue Levels of OrganizationedilbertoNo ratings yet
- Notes On The Phylum PORIFERADocument20 pagesNotes On The Phylum PORIFERAnoblefx100% (1)
- Phylum PoriferaDocument13 pagesPhylum PoriferaKennet CruzNo ratings yet
- Aquatic Book PDFDocument105 pagesAquatic Book PDFMohamed TharwatNo ratings yet
- SpongesDocument29 pagesSpongesDea PramestaNo ratings yet
- Porifera ActivityDocument35 pagesPorifera ActivityNicole De VegaNo ratings yet
- P2C2 Group 3 Lab Report - Protozoa and PoriferaDocument7 pagesP2C2 Group 3 Lab Report - Protozoa and PoriferaAngela D.No ratings yet
- Test Bank For Marketing An Introduction Updated 6Th Canadian Edition Gary Armstrong Philip Kotler Valerie Trifts Lilly Anne Buchwitz Full Chapter PDFDocument24 pagesTest Bank For Marketing An Introduction Updated 6Th Canadian Edition Gary Armstrong Philip Kotler Valerie Trifts Lilly Anne Buchwitz Full Chapter PDFjose.hanson128100% (17)
- Classification of PoriferaDocument5 pagesClassification of Poriferamanojitchatterjee2007No ratings yet
- Prac - 02 Porifera - CtenophoraDocument2 pagesPrac - 02 Porifera - Ctenophorakaytro clashNo ratings yet
- Labporifera 2012Document8 pagesLabporifera 2012PETRO KEWENo ratings yet
- Biodiversity of Shallow-Water Sponges Porifera inDocument19 pagesBiodiversity of Shallow-Water Sponges Porifera inHidayah MushlihahNo ratings yet
- BZ Lab 5Document8 pagesBZ Lab 5Alexa Jean D. HonrejasNo ratings yet
- Leidy 1855Document20 pagesLeidy 1855Afonso Costa Lucas PrestesNo ratings yet
- The Three Grades of Metazoan Animals: Kingdom: AnimaliaDocument52 pagesThe Three Grades of Metazoan Animals: Kingdom: AnimaliaTesfaye SimeNo ratings yet























































































Download as doc, pdf, or txt
You might also like
- Fundamentals of Invertebrate Palaeontology MacrofossilsDocument411 pagesFundamentals of Invertebrate Palaeontology MacrofossilsLuthfan100% (2)
- BiologyDocument140 pagesBiologyZandile MuwaloNo ratings yet
- Chapter 4 - Tissues Glands and MembranesDocument49 pagesChapter 4 - Tissues Glands and MembranesNess G. Mamasabulod100% (1)
- Sponges and PlacozoansDocument30 pagesSponges and Placozoansdayahfarday100% (1)
- The Arthropods: Blueprint For SuccessDocument68 pagesThe Arthropods: Blueprint For SuccessKlym CaballasNo ratings yet
- Phylum Porifera, Characteristics and Class ExamplesDocument4 pagesPhylum Porifera, Characteristics and Class ExamplesRobert BarrettoNo ratings yet
- Phylum PoriferaDocument18 pagesPhylum Poriferagonoles81No ratings yet
- Chapter6-Phylum Porifera NotesDocument4 pagesChapter6-Phylum Porifera Notesapi-195601294No ratings yet
- Classification of ProtozoaDocument1 pageClassification of ProtozoaSalil ShauNo ratings yet
- Biology PDFDocument344 pagesBiology PDFUrmilaAnantNo ratings yet
- Tissues NotesDocument7 pagesTissues Noteskapil100% (1)
- Lab Practical 1Document21 pagesLab Practical 1randy_sanchez_10No ratings yet
- 11.2 Domain Bacteria and ArchaeaDocument39 pages11.2 Domain Bacteria and ArchaeaZkdlin SpaceNo ratings yet
- Plant and Animal CellDocument7 pagesPlant and Animal CellHera Lopez100% (1)
- Frog Lab 1Document4 pagesFrog Lab 1api-296981910No ratings yet
- PoriferaDocument39 pagesPoriferaabhishek.ajmaniNo ratings yet
- Porifera - EvolucionDocument9 pagesPorifera - EvolucionJuan David HPNo ratings yet
- Histo Trans Lec 3 1 PDFDocument8 pagesHisto Trans Lec 3 1 PDFBreville SageNo ratings yet
- Types of BlastulaDocument3 pagesTypes of BlastulaDeepanshu Saini80% (5)
- Lecture - Microbial Diversity The Eukaryotic Microbes - Chapter 4 - BSEDDocument5 pagesLecture - Microbial Diversity The Eukaryotic Microbes - Chapter 4 - BSEDMaden betoNo ratings yet
- Exercise 1 The Cell 1Document6 pagesExercise 1 The Cell 1MJMadlangbayan100% (1)
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Phylum Porifera NotesDocument2 pagesPhylum Porifera Notesapi-24708413675% (4)
- A. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItDocument9 pagesA. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItFrancis100% (1)
- Echinostoma IlocanumDocument3 pagesEchinostoma IlocanumChristie Raye NapuliNo ratings yet
- Phylum EchiuraDocument22 pagesPhylum EchiuraEthyl50% (2)
- Homeostasis and The Excretory SystemDocument21 pagesHomeostasis and The Excretory Systemmark smithNo ratings yet
- Stem Cells in HumansDocument4 pagesStem Cells in HumansSana NainaNo ratings yet
- General Classification (All Phylum)Document16 pagesGeneral Classification (All Phylum)kiran PoudelNo ratings yet
- Phylum Nematoda 13Document132 pagesPhylum Nematoda 13Iseth ISeth100% (1)
- CHAPTER 4 BOTANY - Review Questions and AnswersDocument7 pagesCHAPTER 4 BOTANY - Review Questions and AnswersPam GarciaNo ratings yet
- AnimaldistributioniiiDocument22 pagesAnimaldistributioniiiManoo BiliNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- 11.5 Domain Eukarya: Kingdom PlantaeDocument34 pages11.5 Domain Eukarya: Kingdom PlantaeMuhammad HusniNo ratings yet
- ProtozoansDocument2 pagesProtozoansJonathanPayongayongNo ratings yet
- Animal Nutrition: AP BiologyDocument34 pagesAnimal Nutrition: AP BiologyDoods Galdo100% (1)
- CBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsDocument10 pagesCBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsPriyanshu BhadanaNo ratings yet
- Intestinal and Commensal AmoebaDocument9 pagesIntestinal and Commensal AmoebaFuture TrekingNo ratings yet
- Venn Diagram On Osmosis and DiffusionDocument1 pageVenn Diagram On Osmosis and DiffusioncolacastellNo ratings yet
- Plant CellDocument3 pagesPlant CellSUZAIMAHNo ratings yet
- AMOEBADocument74 pagesAMOEBAhuyenthanh1807No ratings yet
- Cell Structure Function and PropertiesDocument10 pagesCell Structure Function and PropertiesWanda JohnNo ratings yet
- The Primary and Secondary Growth of PlantDocument31 pagesThe Primary and Secondary Growth of PlantNOFAIZAH RAMLINo ratings yet
- Animal Kingdom: Basis of ClassificationDocument15 pagesAnimal Kingdom: Basis of ClassificationChaitali Deshmukh100% (1)
- Cell - Histology Trans Part 1&2Document6 pagesCell - Histology Trans Part 1&2Mark AbrazaldoNo ratings yet
- The Species Concept-1Document5 pagesThe Species Concept-1O me khanNo ratings yet
- Activity 9 Phylum NematodaDocument18 pagesActivity 9 Phylum Nematodanicole syNo ratings yet
- Endocrine SystemDocument27 pagesEndocrine Systemcardos cherry100% (1)
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Histology of Gastrointestinal TractDocument46 pagesHistology of Gastrointestinal Tractandreas kevinNo ratings yet
- Animal Tissues 1Document45 pagesAnimal Tissues 1sirajNo ratings yet
- S.3 Biology Vol. 4 Excretion and HomeostasisDocument13 pagesS.3 Biology Vol. 4 Excretion and HomeostasisFrancheli BautistaNo ratings yet
- Plant KingdomDocument16 pagesPlant Kingdomshriyansh singhaniaNo ratings yet
- 3 ClearingDocument3 pages3 ClearingAngel RamosNo ratings yet
- Leukocytes, Granulocytes, The Monocyte-Macrophage System, and InflammationDocument10 pagesLeukocytes, Granulocytes, The Monocyte-Macrophage System, and InflammationShi no MeNo ratings yet
- Chapter 9 The Immune System NotesDocument8 pagesChapter 9 The Immune System NotesFelix LeNo ratings yet
- 55 16saczo1 2020121910192848Document30 pages55 16saczo1 2020121910192848Dandy H HerkoNo ratings yet
- BSC Part I Canal System in PoriferaDocument6 pagesBSC Part I Canal System in PoriferaSriram MNo ratings yet
- 157 18ZC101 2020120608325782Document18 pages157 18ZC101 2020120608325782pm102382No ratings yet
- PoriferaDocument33 pagesPoriferaRushikesh pawar100% (1)
- Porifera ReviewerDocument8 pagesPorifera ReviewerCarlo MendozaNo ratings yet
- Practical Guide Invertebrates PDFDocument61 pagesPractical Guide Invertebrates PDFMOUSTAFA ALEMAMNo ratings yet
- Guia Visual de Organismos de Aguas Profundas Del Caribe Colombiano / Visual Guide of Deep Sea Organisms of The Colombian CaribbeanDocument99 pagesGuia Visual de Organismos de Aguas Profundas Del Caribe Colombiano / Visual Guide of Deep Sea Organisms of The Colombian CaribbeanAlejandro GarciaNo ratings yet
- Honesty Is The Best Policy EssayDocument8 pagesHonesty Is The Best Policy Essaysmgcjvwhd100% (2)
- Feart 10 914319Document12 pagesFeart 10 914319Satrio Hani SamudraNo ratings yet
- SpongesDocument16 pagesSpongesKazuki Fuchoin100% (1)
- PoriferaDocument33 pagesPoriferaRushikesh pawar100% (1)
- 1st August (Porifera)Document7 pages1st August (Porifera)tayyabuetlhrNo ratings yet
- Chapter 9 - ZoologyDocument3 pagesChapter 9 - ZoologyAngel MerilloNo ratings yet
- Phylum Cnidaria and Porifera Group 1Document32 pagesPhylum Cnidaria and Porifera Group 1JulesNo ratings yet
- PoriferaDocument11 pagesPoriferaJohn AdamNo ratings yet
- Sponge ColoringDocument2 pagesSponge ColoringMathias CronqvistNo ratings yet
- Practical Manual in Zoology: Dr. A. AmsathDocument323 pagesPractical Manual in Zoology: Dr. A. AmsathAnnapoorna SagarNo ratings yet
- The Fauna Associated With Outer Shelf and Upper Slope SpongesDocument22 pagesThe Fauna Associated With Outer Shelf and Upper Slope SpongesPedro Henrique ClerierNo ratings yet
- Porififera and CnidariaDocument6 pagesPorififera and CnidariaFlorence PalomarNo ratings yet
- Module 8 - Multi-Cellular and Tissue Levels of OrganizationDocument25 pagesModule 8 - Multi-Cellular and Tissue Levels of OrganizationedilbertoNo ratings yet
- Notes On The Phylum PORIFERADocument20 pagesNotes On The Phylum PORIFERAnoblefx100% (1)
- Phylum PoriferaDocument13 pagesPhylum PoriferaKennet CruzNo ratings yet
- Aquatic Book PDFDocument105 pagesAquatic Book PDFMohamed TharwatNo ratings yet
- SpongesDocument29 pagesSpongesDea PramestaNo ratings yet
- Porifera ActivityDocument35 pagesPorifera ActivityNicole De VegaNo ratings yet
- P2C2 Group 3 Lab Report - Protozoa and PoriferaDocument7 pagesP2C2 Group 3 Lab Report - Protozoa and PoriferaAngela D.No ratings yet
- Test Bank For Marketing An Introduction Updated 6Th Canadian Edition Gary Armstrong Philip Kotler Valerie Trifts Lilly Anne Buchwitz Full Chapter PDFDocument24 pagesTest Bank For Marketing An Introduction Updated 6Th Canadian Edition Gary Armstrong Philip Kotler Valerie Trifts Lilly Anne Buchwitz Full Chapter PDFjose.hanson128100% (17)
- Classification of PoriferaDocument5 pagesClassification of Poriferamanojitchatterjee2007No ratings yet
- Prac - 02 Porifera - CtenophoraDocument2 pagesPrac - 02 Porifera - Ctenophorakaytro clashNo ratings yet
- Labporifera 2012Document8 pagesLabporifera 2012PETRO KEWENo ratings yet
- Biodiversity of Shallow-Water Sponges Porifera inDocument19 pagesBiodiversity of Shallow-Water Sponges Porifera inHidayah MushlihahNo ratings yet
- BZ Lab 5Document8 pagesBZ Lab 5Alexa Jean D. HonrejasNo ratings yet
- Leidy 1855Document20 pagesLeidy 1855Afonso Costa Lucas PrestesNo ratings yet
- The Three Grades of Metazoan Animals: Kingdom: AnimaliaDocument52 pagesThe Three Grades of Metazoan Animals: Kingdom: AnimaliaTesfaye SimeNo ratings yet