Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- DRYPIX Smart Reference GuideDocument80 pagesDRYPIX Smart Reference GuideHugo79% (33)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- NFPA 13 PRESSURE TEST PROCEDURES (Fire Pro)Document2 pagesNFPA 13 PRESSURE TEST PROCEDURES (Fire Pro)Mechanical Engineer33% (3)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Complete Jason Statham WorkoutDocument11 pagesThe Complete Jason Statham WorkoutCristi OcuNo ratings yet
- Butterfly ValveDocument24 pagesButterfly ValveJoy Arief JanuarNo ratings yet
- Aaron VanneyDocument48 pagesAaron VanneyIvan KelamNo ratings yet
- K K A C D W K P S K T: Eteraturan Unjungan Ntenatal ARE I Ilayah Erja Uskesmas Lawi Abupaten EgalDocument6 pagesK K A C D W K P S K T: Eteraturan Unjungan Ntenatal ARE I Ilayah Erja Uskesmas Lawi Abupaten EgalSelly MonicaNo ratings yet
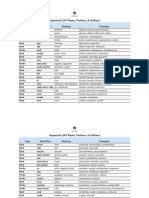
- Most Common Roots, Prefixes & Suffixes - Google SheetsDocument8 pagesMost Common Roots, Prefixes & Suffixes - Google Sheetsnischal05pNo ratings yet
- 52369.Đề ôn thi lớp 10Document5 pages52369.Đề ôn thi lớp 10Phương LinhNo ratings yet
- Annex Q - Project Completion ReportDocument2 pagesAnnex Q - Project Completion ReportPBT apstNo ratings yet
- Risk Management in Commercial BanksDocument37 pagesRisk Management in Commercial BanksRavi DepaniNo ratings yet
- Tle 7 - Q1.W1Document2 pagesTle 7 - Q1.W1Pia DelaCruz100% (2)
- An Aspen Plus® Tool For Simulation of PDFDocument16 pagesAn Aspen Plus® Tool For Simulation of PDFMahmud Arman KibriaNo ratings yet
- Stepladder Plating Following Lower Subapical and Geniop - 2022 - Advances in OraDocument2 pagesStepladder Plating Following Lower Subapical and Geniop - 2022 - Advances in OraГне ДзжNo ratings yet
- EMS14 EMS Products Only Catalogue 2022 V17 FINAL ScreenDocument20 pagesEMS14 EMS Products Only Catalogue 2022 V17 FINAL ScreenAbdallah MohamedNo ratings yet
- Plpro 2Document1 pagePlpro 2Amal VettomNo ratings yet
- Majdanek, The Concetration Camp of LublinDocument72 pagesMajdanek, The Concetration Camp of Lublinheyyou100% (3)
- Estimation of The Vitamin D Content of Foods For The Assessment of Dietary IntakeDocument1 pageEstimation of The Vitamin D Content of Foods For The Assessment of Dietary IntakeNancy Kosasih 2No ratings yet
- Phytochemical Screening of Certain Medicinal Plants of Mizoram India and Their Folklore Use 2376 0214.1000136Document9 pagesPhytochemical Screening of Certain Medicinal Plants of Mizoram India and Their Folklore Use 2376 0214.1000136radivyenghuNo ratings yet
- Sid97 3eDocument39 pagesSid97 3eJonatan BernalNo ratings yet
- Gr-4 Bridge Course Day 3Document17 pagesGr-4 Bridge Course Day 3Asif KNo ratings yet
- Safety Inspection Vs Safety AuditDocument10 pagesSafety Inspection Vs Safety Auditاحمد عبداللاNo ratings yet
- Approach To Patient With Altered SensoriumDocument4 pagesApproach To Patient With Altered SensoriumMiguel Cuevas DolotNo ratings yet
- StyrofoamDocument9 pagesStyrofoamapi-353356333No ratings yet
- NEET UG Biology Molecular Basis of InheritanceDocument18 pagesNEET UG Biology Molecular Basis of InheritanceNárēsh Yadav GäddēNo ratings yet
- MS P2 CompilationDocument451 pagesMS P2 CompilationJe KirsteneNo ratings yet
- Report On Degraded Ecosystem Pashan LakeDocument13 pagesReport On Degraded Ecosystem Pashan LakeArash AhmadNo ratings yet
- Fireman's Rule and Its Extension To EMT'sDocument7 pagesFireman's Rule and Its Extension To EMT'sShahin DamouiNo ratings yet
- Ds About Magnesium OilDocument2 pagesDs About Magnesium OilMark Dingle100% (1)
- Reactions of Synthetic ImportanceDocument28 pagesReactions of Synthetic ImportanceRx Nadeem ChhipaNo ratings yet
- LEVOXIN eDocument1 pageLEVOXIN ejim_corbet1211No ratings yet