
30 Vijaykumar Etal
30 Vijaykumar Etal
You might also like
- Bamboos of India WebDocument354 pagesBamboos of India WebChristian Gauss100% (1)
- GR 10 Biology - Tissues Organs and System of Living ThingDocument28 pagesGR 10 Biology - Tissues Organs and System of Living Thingapi-301141568No ratings yet
- 100 Concepts of Developmental and Gross AnatomyDocument246 pages100 Concepts of Developmental and Gross AnatomyAbd Samad100% (1)
- Contractilidad Bombardini2005Document22 pagesContractilidad Bombardini2005Carlos Navarro alonsoNo ratings yet
- Jurnal 55Document1 pageJurnal 55rizalmantaNo ratings yet
- CalcioDocument31 pagesCalcioAlejandro Londoño AmarilesNo ratings yet
- Sarcoplasmic Reticulum 3Document6 pagesSarcoplasmic Reticulum 3Syaidah umiNo ratings yet
- UpToDate - Anti Arrhythmic Drugs MOADocument31 pagesUpToDate - Anti Arrhythmic Drugs MOA9821699080No ratings yet
- Calcium Sparks: Physiol RevDocument56 pagesCalcium Sparks: Physiol RevFisiologiaUFCGNo ratings yet
- Cardiac Pacing Learning Package 2013Document34 pagesCardiac Pacing Learning Package 2013Chetan Amberkar100% (1)
- 18sodium Calcium ExchangerDocument4 pages18sodium Calcium ExchangerZiedTrikiNo ratings yet
- Advanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Document28 pagesAdvanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Muhammad Mudassar ChudaryNo ratings yet
- Disorders of Heart RhythmsDocument202 pagesDisorders of Heart Rhythmspraveena.pv1234No ratings yet
- Cardiac Action Potential - WikipediaDocument67 pagesCardiac Action Potential - WikipediaMohd. Mudabbir-Ul- IslamNo ratings yet
- Principles of Cardiac ElectrophysiologyDocument98 pagesPrinciples of Cardiac ElectrophysiologyAbnet WondimuNo ratings yet
- Cardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateDocument32 pagesCardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateClaudia AlessandraNo ratings yet
- Jphysiol 2009 181578Document2 pagesJphysiol 2009 181578ProtopaticoNo ratings yet
- Logy Assighnment On Automaticity Rhythmicity AutorecoveredDocument10 pagesLogy Assighnment On Automaticity Rhythmicity AutorecoveredM Asif RiazNo ratings yet
- Cardiac Action PotentialDocument5 pagesCardiac Action PotentialjayNo ratings yet
- Signaling in Muscle ContractionDocument15 pagesSignaling in Muscle ContractionMARTINEZ SANCHEZ JULIAN ELIASNo ratings yet
- CalcioDocument6 pagesCalcioArtur Franck Tenorio AronésNo ratings yet
- Physiology FAQ 2018Document185 pagesPhysiology FAQ 2018kirki pNo ratings yet
- Hypokalemia and Anesthetic Implications: Wong, Patrick Schafer, and John R. SchultzDocument23 pagesHypokalemia and Anesthetic Implications: Wong, Patrick Schafer, and John R. SchultzRidski D. MiruNo ratings yet
- Excitation PrintDocument3 pagesExcitation PrintZoRo4everNo ratings yet
- Heart PhysiolDocument84 pagesHeart PhysiolChintya Ayu ChampakaNo ratings yet
- L-Type Calcium ChannelDocument3 pagesL-Type Calcium ChannelNTA UGC-NETNo ratings yet
- Signaling in Muscle ContractionDocument14 pagesSignaling in Muscle ContractionJosué VelázquezNo ratings yet
- Nerves, Hormones and HomeostasisDocument8 pagesNerves, Hormones and Homeostasisumairas_1No ratings yet
- Oakes-Mitochondria Ca Immune SynapseDocument2 pagesOakes-Mitochondria Ca Immune Synapseiulia andreeaNo ratings yet
- Cytosolic Calcium Homeostasis in Neurons - Control Systems, Modulation by Reactive Oxygen and Nitrogen Species, and Space and Time FluctuationsDocument52 pagesCytosolic Calcium Homeostasis in Neurons - Control Systems, Modulation by Reactive Oxygen and Nitrogen Species, and Space and Time Fluctuationskj185No ratings yet
- Channelopathies: Ion Channel Defects Linked To Heritable Clinical DisordersDocument12 pagesChannelopathies: Ion Channel Defects Linked To Heritable Clinical Disorderscote_cote6951No ratings yet
- Llinas PNASDocument6 pagesLlinas PNASjhrubioNo ratings yet
- Molecular Basis of Cardiac Action Potential RepolarizationDocument6 pagesMolecular Basis of Cardiac Action Potential RepolarizationJuan Jose Valladares SanchezNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologySteve FerisNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologyTamjid Khan100% (2)
- Arrythmia by AbnetDocument509 pagesArrythmia by AbnetAbnet WondimuNo ratings yet
- Calcium in BiologyDocument8 pagesCalcium in BiologyomNo ratings yet
- Review Article Crosstalk Between Calcium and ROS in Pathophysiological ConditionsDocument19 pagesReview Article Crosstalk Between Calcium and ROS in Pathophysiological ConditionsbetiNo ratings yet
- Action PotentialDocument2 pagesAction PotentialMian Talha KamalNo ratings yet
- Cardiac Muscle Physiology: P. Szedlak, D.S. Steele and P.M. HopkinsDocument8 pagesCardiac Muscle Physiology: P. Szedlak, D.S. Steele and P.M. Hopkinsmastmulla921No ratings yet
- Tissue To Organisms/Physiology & PharmacologyDocument6 pagesTissue To Organisms/Physiology & PharmacologyStudent LifeNo ratings yet
- Praktikum Faal-Blok14-3BDocument18 pagesPraktikum Faal-Blok14-3BKelompok 5A Semester 4No ratings yet
- Exercise-Induced Rhabdomyolysis Mechanisms and PreventionDocument11 pagesExercise-Induced Rhabdomyolysis Mechanisms and PreventionMarlene PgNo ratings yet
- Cellular Biology of EpileptogenesisDocument10 pagesCellular Biology of EpileptogenesisandreacamachogonNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsleandronascciNo ratings yet
- QSAR Studies On Calcium Channel Blockers: Satya P. GuptaDocument39 pagesQSAR Studies On Calcium Channel Blockers: Satya P. GuptaOana FloreanNo ratings yet
- UNIT 9 NotesDocument5 pagesUNIT 9 NotesMariaNo ratings yet
- The Journal of Physiology - 2008 - Berridge - Smooth Muscle Cell Calcium Activation MechanismsDocument15 pagesThe Journal of Physiology - 2008 - Berridge - Smooth Muscle Cell Calcium Activation MechanismsNoumanNo ratings yet
- Human Physiology Assignment 01Document9 pagesHuman Physiology Assignment 01Evaristo MK VincentNo ratings yet
- The Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesDocument30 pagesThe Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesKhel VincentNo ratings yet
- Pi Is 0022282817303115Document11 pagesPi Is 0022282817303115EdgarNo ratings yet
- Neurochem FinalDocument20 pagesNeurochem Finalelifulku.a1No ratings yet
- 1.bomba de Na - KDocument22 pages1.bomba de Na - KJosé DiegoNo ratings yet
- Potassium Channels ThesisDocument7 pagesPotassium Channels Thesisbk156rhq100% (2)
- Computer SimulationDocument29 pagesComputer SimulationmrtiborhornyikNo ratings yet
- Skeletal Muscle Physiology PDFDocument6 pagesSkeletal Muscle Physiology PDFAstri Ggamjong Xiao LuNo ratings yet
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Cardiac PDFDocument49 pagesCardiac PDFViswani pillapalyamNo ratings yet
- Cardiac Action PotentialDocument41 pagesCardiac Action PotentialRadijska Postaja KoprivnicaNo ratings yet
- Advanced Cell BiologyDocument14 pagesAdvanced Cell BiologyUmair AneesNo ratings yet
- Learning Objectives: Chapter 4: Electrophysiology of The HeartDocument28 pagesLearning Objectives: Chapter 4: Electrophysiology of The HeartLaura PucheNo ratings yet
- DISCUDocument23 pagesDISCUpvsvrNo ratings yet
- EKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXFrom EverandEKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXRating: 5 out of 5 stars5/5 (2)
- Ijmrhs Vol 4 Issue 2Document219 pagesIjmrhs Vol 4 Issue 2editorijmrhsNo ratings yet
- Ijmrhs Vol 4 Issue 4Document193 pagesIjmrhs Vol 4 Issue 4editorijmrhs0% (1)
- Ijmrhs Vol 4 Issue 3Document263 pagesIjmrhs Vol 4 Issue 3editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 4Document294 pagesIjmrhs Vol 3 Issue 4editorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 4Document321 pagesIjmrhs Vol 2 Issue 4editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 2Document281 pagesIjmrhs Vol 3 Issue 2editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 1Document228 pagesIjmrhs Vol 3 Issue 1editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 3Document271 pagesIjmrhs Vol 3 Issue 3editorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 3Document399 pagesIjmrhs Vol 2 Issue 3editorijmrhs100% (1)
- Ijmrhs Vol 1 Issue 1Document257 pagesIjmrhs Vol 1 Issue 1editorijmrhsNo ratings yet
- 48 MakrandDocument2 pages48 MakrandeditorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 1Document110 pagesIjmrhs Vol 2 Issue 1editorijmrhs100% (1)
- Ijmrhs Vol 2 Issue 2Document197 pagesIjmrhs Vol 2 Issue 2editorijmrhsNo ratings yet
- 35krishnasamy EtalDocument1 page35krishnasamy EtaleditorijmrhsNo ratings yet
- 47serban Turliuc EtalDocument4 pages47serban Turliuc EtaleditorijmrhsNo ratings yet
- Recurrent Cornual Ectopic Pregnancy - A Case Report: Article InfoDocument2 pagesRecurrent Cornual Ectopic Pregnancy - A Case Report: Article InfoeditorijmrhsNo ratings yet
- 45mohit EtalDocument4 pages45mohit EtaleditorijmrhsNo ratings yet
- Pernicious Anemia in Young: A Case Report With Review of LiteratureDocument5 pagesPernicious Anemia in Young: A Case Report With Review of LiteratureeditorijmrhsNo ratings yet
- Williams-Campbell Syndrome-A Rare Entity of Congenital Bronchiectasis: A Case Report in AdultDocument3 pagesWilliams-Campbell Syndrome-A Rare Entity of Congenital Bronchiectasis: A Case Report in AdulteditorijmrhsNo ratings yet
- 36rashmipal EtalDocument6 pages36rashmipal EtaleditorijmrhsNo ratings yet
- 37poflee EtalDocument3 pages37poflee EtaleditorijmrhsNo ratings yet
- 38vaishnavi EtalDocument3 pages38vaishnavi EtaleditorijmrhsNo ratings yet
- 41anurag EtalDocument2 pages41anurag EtaleditorijmrhsNo ratings yet
- 40vedant EtalDocument4 pages40vedant EtaleditorijmrhsNo ratings yet
- 31tushar EtalDocument4 pages31tushar EtaleditorijmrhsNo ratings yet
- 34tupe EtalDocument5 pages34tupe EtaleditorijmrhsNo ratings yet
- 33 Prabu RamDocument5 pages33 Prabu RameditorijmrhsNo ratings yet
- 28nnadi EtalDocument4 pages28nnadi EtaleditorijmrhsNo ratings yet
- Introduction (Recovered)Document37 pagesIntroduction (Recovered)muhamedshuaibbhaiNo ratings yet
- Anatomy of The Nervous SystemDocument85 pagesAnatomy of The Nervous SystemjoelNo ratings yet
- Johnson Et Al.2006. Felidae Phylogeny PDFDocument6 pagesJohnson Et Al.2006. Felidae Phylogeny PDFNydia JGNo ratings yet
- Bamboo Species Source List No. 36 2017-2018Document56 pagesBamboo Species Source List No. 36 2017-2018André MichaelNo ratings yet
- Nettle FibersDocument7 pagesNettle Fibersawais123142541254125No ratings yet
- Arthroscopy of The Temporomandibular JointDocument44 pagesArthroscopy of The Temporomandibular JointDencyclopediaNo ratings yet
- Classification of OrganismDocument35 pagesClassification of OrganismJnll JstnNo ratings yet
- SIPDocument20 pagesSIPLuiiiNo ratings yet
- Grade 11 Q2 Module 6 Week 6Document11 pagesGrade 11 Q2 Module 6 Week 6Shiela MaeNo ratings yet
- Grassland Landscape ManagementDocument22 pagesGrassland Landscape ManagementAr Deepika RajaNo ratings yet
- The Fundamental Unit of LifeDocument16 pagesThe Fundamental Unit of LifeSriram RNo ratings yet
- Type-I Collagen Amino Acid Glycine Collagen Triple HelixDocument8 pagesType-I Collagen Amino Acid Glycine Collagen Triple Helixkyle swayNo ratings yet
- Hill Stream AdaptationDocument5 pagesHill Stream AdaptationAdarsh SenNo ratings yet
- Anxiety Lesson WorksheetDocument4 pagesAnxiety Lesson WorksheetAmisha VastaniNo ratings yet
- Recommended Procedures For Sample Collection Preservation and Shipping-D.senneDocument48 pagesRecommended Procedures For Sample Collection Preservation and Shipping-D.senneAndersonNo ratings yet
- Skin Conditions Worksheet 2017Document3 pagesSkin Conditions Worksheet 2017Tamia WatsonNo ratings yet
- AnthDocument18 pagesAnthAnurag ShuklaNo ratings yet
- Evolution of AnuraDocument18 pagesEvolution of AnuraSan Nguyễn ViếtNo ratings yet
- Nat 5 Biology SQ App 2019Document48 pagesNat 5 Biology SQ App 2019muntazNo ratings yet
- New English Sba 2018Document15 pagesNew English Sba 2018ashawni patterson88% (8)
- 2024 Msyi Final ProgramDocument12 pages2024 Msyi Final Programapi-654587216No ratings yet
- Result Oriented Management (ROM) - 2Document7 pagesResult Oriented Management (ROM) - 2baguma100% (1)
- Biology and Physiology of Saline PlantsDocument32 pagesBiology and Physiology of Saline PlantsNguyễn Lê Quốc ThiNo ratings yet
- Scientific Aspects in Ramcharit ManasDocument52 pagesScientific Aspects in Ramcharit ManasRam Kumar Kuril100% (9)
- 16625.Hsl4207 Microbiology (Parasitology & Mycology) CH 13Document4 pages16625.Hsl4207 Microbiology (Parasitology & Mycology) CH 13Tarun AroraNo ratings yet
- Assessment Diagnosis Scientific Reason Planning Intervention Rational E EvaluationDocument2 pagesAssessment Diagnosis Scientific Reason Planning Intervention Rational E EvaluationCamille VirayNo ratings yet
- Supplement Guide SleepDocument39 pagesSupplement Guide SleepinfoNo ratings yet
























































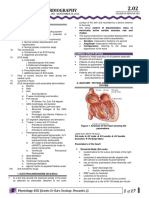





























































DOI: 10.5958/2319-5886.2015.00077.
International Journal of Medical Research
&
Health Sciences
www.ijmrhs.com
Volume 4 Issue 2
Coden: IJMRHS
th
Received: 20 Jan 2015
Revised: 28th Feb 2015
Review article
Copyright @2015 ISSN: 2319-5886
Accepted: 19th Mar 2015
EVOLUTION OF AUTOMATICITY OF HEART PACEMAKER STUDIED FROM A THEORETICAL
PERSPECTIVE
3
Vijay Kumar Konuri1, Mohammed Abdul Hannan Hazari2, Ravi Kumar K , Chandrasekhar M ,
5
Ambareesha K , Ram Reddy B
1
Associate Professor, Department of Anatomy, All India Institute of Medical Sciences, Raipur, Chhattisgarh,
India
2
Associate Professor, Department of Physiology, Deccan College of Medical Sciences, Kanchanbagh, Hyderabad,
Telangana, India
3
Assistant Professor of Anatomy, Govt Medical College, Jagdalpur, Chhattisgarh
Professor and HOD, 5Tutor, Department of Physiology, Meenakshi Medical College, Kanchipuram,
Tamil Nadu
4
Professor & HOD, Department of Physiology, Apollo Institute of Medical Sciences and Research, Jubilee Hills,
Hyderabad, Telangana, India.
*Corresponding author email: vkkonuri@gmail.com
ABSTRACT
The pacemaker of the mammalian heart had developed a robust and yet a flexible system in the course of
evolution whose function is based on multiple interactions at the sub-cellular, cellular and finally at the tissue
level. These, in turn, should respond to extrinsic signals. Cardiac action potentials were explained for a long time
based on the changes that occur at the cell surface. New hypothesis was put forward at the turn of the century that
pointed to the role of intracellular calcium clock. Discovery of ryanodine receptors, fluorescence labeling
techniques, confocal imaging and finally computer modeling of physiological processes had brought about a
noticeable change that allowed development of a new concept of pacemaker automaticity. Reviewing all these
developments we hereby put forward a few theoretical formulations that can turn out to be new instruments in
advancing our knowledge of cardiac physiology. We had theorized that cardiac muscle is an emergent property of
smooth muscle in the course of evolution, and that pacemaker activity of the cardiac muscle underwent a phase
transition that finally led to the evolution of a structural pacemaker.
Keywords: Heart, Pacemaker, Automaticity, Evolution
INTRODUCTION
The sino-atrial node (SAN) pacemaker cells produce
billions of incessant and uninterrupted beats in the
course of the life time of an individual. It is evident
that the pacemaker of the mammalian heart has
developed a robust and yet a flexible system in the
course of evolution [1]. Robustness indicates the failsafe properties and flexibility signifies the
adaptability to changes in the demands made on it.
The pacemaker function is based on multiple
interactions at the level of sub-cellular, cellular and
finally at the level of tissue architecture, which in turn
should react to extrinsic signals like stretch, electrical
and chemical signals that act on the cell surface
receptors.
The generation of action potentials in the
myocardium was explained for a long time,
predominantly, basing on the changes that occur on
the cell surface and its ion channels [2]. However, the
417
Vijaykumar et al.,
Int J Med Res Health Sci. 2015;4(2):417-421
turn of the century has brought new evidence pointing
to the role of an intracellular clock. This turned out to
be the cyclical rhythm of cytosolic Ca2+
concentration. Sarcoplasmic reticulum (SR) has
proved to be having the capacity to operate another
physiologic clock of calcium cycles[3]. Recent
developments in experimentation like confocal
imaging have revealed the presence of multiple
spontaneous, rhythmic, local calcium releases[4].
These tightly regulated processes begin to occur
beneath the cell surface during the later part of
diastolic depolarization. This activates the Na+-Ca2+
exchanger that causes an explosive increase in
diastolic depolarization and leads to the activation of
L-type calcium channels[5].
Fig 1. Interplay between intracellular and membrane
surface Ca2+ clock.
The existing understanding of cardiac action
potentials
The explanation of action potentials in the working
myocardium is as follows. Phase 0 or the rapid
depolarization results from the opening of the fast
Na+ channels. Phase 1 of the action potential starts
the repolarization process and is attributed to the
closing of Na+ channels and inward movement of Clions. Phase 2, the so-called "plateau" phase of the
action potential results from several mechanisms but
could be mainly because of slow inward movement of
Ca2+ and Na+. Phase 3 involves relative rapid
repolarization, commencing with inactivation of slow
Ca2+ and Na+ channels and rapid outward movement
of K+ ions. Phase 4 restores the ionic composition
back to the resting state by Na+-K+ ATPase which
pump Na+ ions out and K+ ions inside the cell[6].
Membrane
potential
of
pacemaker
cells
spontaneously declines to the firing level and is
known as prepotential or pacemaker potential which
triggers the next action potential. At the peak of each
action potential, conductance of potassium (IK+)
begins and repolarization occurs. IK+ then declines
and the membrane potential reaches slight
hyperpolarization. At this instance an "h" or "f"
channel which allows both Na+ and K+ is activated.
As conductance through "h" channel (Ih) increases,
the membrane begins to depolarize forming the initial
part of the prepotential. T-type Ca2+ channels then
open and its conductance (ICa2+T) completes the
prepotential. At this juncture L-type Ca2+ channels
open and ICa2+L produce action potential.
The measurements of calcium concentrations
The finding that oscillations in Ca2+ concentrations
were inhibited by calcium channel blockers (CCBs)
had brought to the fore the idea that Ca2+
concentration represents a two-way interaction
between the intracellular Ca2+ stores and the
membrane surface potential changes[7]. But due to the
rapidity of changes; spontaneous, localized
oscillations of the calcium clock could not be
measured within the individual SA nodal cells and
hence the concept that initiators of the normal
automaticity of pacemaker cells are internal calcium
oscillations could not be established.
The membrane surface processes was disconnected
from that of intracellular oscillations, for a long time,
by the employment of Ca2+ overload conditions to
voltage clamp studies[8]. Studies have gradually
demonstrated that the intracellular oscillations could,
in fact, produce spontaneous membrane currents. It is
now considered that the intracellular oscillations
involve the cycling of Ca2+ ions between SR and
cytosol[9].
Discovery of ryanodine receptors
Then came the discovery that the drug named
ryanodine can have a profound negative chronotropic
effect on the automaticity of cardiac pacemaker cells.
By studying the effect of ryanodine on the contour of
the action potential it was suggested that Ca2+
released from the SR contributes to diastolic
depolarization[10].
It is now possible to measure the intracellular calcium
levels in spontaneously firing pacemaker cells of
SAN which made it clear that each spontaneous
action potential evokes a calcium gradient in the
cytosol and that the influx of calcium through L-type
calcium channels affects the calcium loading of the
418
Vijaykumar et al.,
Int J Med Res Health Sci. 2015;4(2):417-421
SR. Intracellular buffering of calcium has the potency
to block the generation of spontaneous action
potentials[11]. This constitutes a strong evidence in
favor of the idea that normal automaticity of
pacemaker cells is strongly linked with the dynamics
of intracellular calcium.
Modern techniques
Fluorescence imaging of intracellular calcium is
made possible in the last decade that enabled to
document not only the global transients of cytosolic
calcium but also of many localized calcium releases
beneath the cell surface during late diastolic
depolarization[12]. Such local calcium releases are
observed in SA nodal cells in the absence of changes
in the membrane potentials, i.e. in voltage clamped
SA node pacemaker cells. There is evidence that local
Ca2+ release from the sarcoplasmic reticulation (Ca2+
sparks) occurs during the prepotential. Local calcium
releases (LCRs) during the late diastolic
depolarization begin to boil and then explode into an
action potential[13].
A tiny change in the current to the degree of 3 pA is
enough to explode into an action potential during the
critical diastolic depolarization phase of rabbit SA
node cells. Although the individual local release of
calcium during the diastolic depolarization of
pacemaker cells is relatively small and stochastic in
nature, the synchronized and cumulative effects of
LCRs imparts and impacts the rising phase of
diastolic depolarization leading to the next action
potential. A failure to generate an exponential phase
in diastolic depolarization is the consequence of a
failure to generate diastolic INCX[14].
Structurally SA node is heterogeneous
Till now we discussed the mechanisms of
automaticity and spontaneous calcium cycles in
individual pacemaker cells. But cardiac pacemaker
function cannot be understood completely by the
study of the intrinsic properties of the pacemaker
cells. Advanced histological studies had revealed that
SA node is a highly heterogeneous structure with
small pacemaker cells predominantly located in the
centre[15]. The SA nodal tissue is characterized by
complex cell-to-cell interactions in generating the
highly robust impulses with a fail-safe mechanism[16].
The function of the pacemaker tissue within the SA
node is determined by the intrinsic properties of
individual cells that are being modulated by several
factors of the local environment within the SA
node[17]. The pacemaker tissue is at the same time
being influenced by extrinsic modulators that include
the electrical and mechanical forces as well as the
autonomic milieu. These modulatory factors are
heterogeneous throughout the SA node which could
explain the differences in the shape of the action
potential curves of different cells in the same
locality[18]. This could be the result of mutual
interaction between the depolarizing charges
generated by individual SA node cells and the
structural properties of the surrounding non-excitable
tissue[19].
Extensive amounts of connective tissue and numerous
fibroblasts occupy from 25% to almost 90% of the
area of SA node[20].
Fig 2: Phase transition and emergence of cardiac muscle from smooth muscle with maintenance of legacy
in some aspects
419
Vijaykumar et al.,
Int J Med Res Health Sci. 2015;4(2):417-421
In addition to bands of connective tissue, the gap
junction proteins are also found to be located in
various types and with varying densities in near
vicinities. The myofilament density decreases from
periphery to the centre of the SA node[21]. The nerve
endings and autonomic receptors on pacemaker cells
are found to be at the highest density in the central
area of SA node[22]. All these structural features make
the SA node a highly heterogeneous structure that can
generate not only flexible but also a robust action
potential[23].Intracellular calcium cycling plays a
major role in the generation of automaticity in
embryonic cardio myocytes and so could be used to
generate stem cell derived spontaneously beating
myoblast cells[24].
Discussion: A theoretical approach is needed to
interpret physiology at an advanced level
We have seen how our concepts of pacemaker
potentials transformed over time, from the more or
less simplified notions that membrane currents
explain everything to developing a more and more
complex picture of mutual entrainment of the
cytosolic and the membrane clocks and of their
different mechanisms in generating automaticity.
We understand the vertebrate circulatory system as a
closed canal system comprised of network of smooth
muscle and other cells, the proximal part of which
has been transformed into cardiac muscle due to
increased circulatory load imposed on the
system[25].This can be visualized as a type of phase
transition during evolution of smooth muscle there by
transforming phase maker activity from a network of
molecules to a network of cells and tissues, which can
justify the existence of special conducting system in
myocardium. So this can be generalized as an
emergent property of smooth muscle. Pacemaker
properties of the smooth muscle is well preserved
but is now regulated by pacemaker system of cardiac
muscle. Even in smooth muscle the initiation of
pacemaker activity is due to the oscillations of
intracellular calcium that are being modulated by
external conditions. It is our endeavor to elucidate
how function is translated into structure through the
alteration of the genetic program.
CONCLUSION
Pacemaker in mammalian heart is the phase shift
transformation of the smooth muscle through cardiac
muscle during the evolutionary process. In the course
of development of highly evolved forms of
organisms, the metabolic necessities of the complex
tissues and organs demanded continuous supply of
nutrients and gases for sustenance of life. Hence, the
smooth muscle underwent structural and functional
adaptations to develop into cardiac muscle which
further acquired autonomy at the expense of losing
the contractility to form pacemaker tissue. This aptly
describes the proverb "Necessity is the mother of
invention".
REFERENCES
1. Lakatta EG, Vinogradova TM, Maltsev VA. The
missing link in the mystery of normal
automaticity of cardiac pacemaker cells. Ann N
Y Acad Sci. 2008; 1123: 4157.
2. Mangoni ME and Nargeot J. Genesis and
regulation of the heart automaticity. Physiol Rev.
2008; 88:919982.
3. Lakatta EG, Vinogradova T, Lyashkov A,
Sirenko S, Zhu W, Ruknudin A, Maltsev VA.
The integration of spontaneous intracellular Ca2+
cycling and surface membrane ion channel
activation entrains normal automaticity in cells of
the hearts pacemaker. Ann N Y Acad Sci. 2006;
1080:178206.
4. Maltsev VA and Lakatta EG. Synergism of
coupled sub-sarcolemmal Ca2+ clocks and
sarcolemmal voltage clocks confers robust and
flexible pacemaker function in a novel pacemaker
cell model. Am J Physiol Heart Circ Physiol.
2009; 296:H594H615.
5. Maltsev VA and Lakatta EG. Dynamic
interactions of an intracellular Ca2+ clock and
membrane ion channel clock underlie robust
initiation and regulation of cardiac pacemaker
function. Cardiovasc Res. 2008; 77:274284.
6. Sanders L, Rakovic S, Lowe M, Mattick PA,
Terrar DA. Fundamental importance of Na-Ca2+
exchange for the pacemaking mechanism in
guinea-pig sino-atrial node. J Physiol. 2006:
571:639649.
7. Vinogradova TM and Lakatta EG. Regulation of
basal and reserve cardiac pacemaker function by
420
Vijaykumar et al.,
Int J Med Res Health Sci. 2015;4(2):417-421
8.
9.
10.
11.
12.
13.
14.
15.
interactions of cAMP mediated PKA-dependent
Ca2+ cycling with surface membrane channels. J
Mol Cell Cardiol. 2009; 47:456 474.
Joung B, Tang L, Maruyama M, Han S, Chen Z,
Stucky M, Jones LR, Fishbein MC, Weiss JN,
Chen PS, Lin SF. Intracellular calcium dynamics
and acceleration of sinus rhythm by betaadrenergic stimulation. Circulation. 2009;
119:788796.
Rigg L, Heath BM, Cui Y, Terrar DA.
Localisation and functional significance of
ryanodine receptors during beta-adrenoceptor
stimulation in the guinea-pig sino-atrial node.
Cardiovasc Res. 2000; 48:254264.
Li J, Qu J, Nathan RD. Ionic basis of ryanodines
negative chronotropic effect on pacemaker cells
isolated from the sinoatrial node. Am J Physiol.
1997; 273:H2481H2489.
Bogdanov KY, Vinogradova TM, Lakatta EG.
Sinoatrial nodal cell ryanodine receptor and Na+Ca2+ exchanger: molecular partners in pacemaker
regulation. Circ Res. 2001; 88:12541258.
Vinogradova TM, Lyashkov AE, Zhu W,
Ruknudin AM, Sirenko S, Yang D, Deo S,
Barlow M, Johnson S, Caffrey JL. Zhou YY,
Xiao RP, Cheng H, Stern MD, Maltsev VA,
Lakatta EG. High basal protein kinase Adependent phosphorylation drives rhythmic
internal Ca2+ store oscillations and spontaneous
beating of cardiac pacemaker cells. Circ Res.
2006; 98:505514.
Maltsev VA and Lakatta EG. Cardiac pacemaker
cell failure with preserved If, ICaL, and IKr: a
lesson about pacemaker function learned from
ischemia-induced bradycardia. J
Mol Cell
Cardiol. 2007; 42:289294.
Vinogradova TM, Zhou YY, Maltsev V,
Lyashkov A, Stern M, Lakatta EG. Rhythmic
ryanodine receptor Ca2+ releases during diastolic
depolarization of sinoatrial pacemaker cells do
not require membrane depolarization. Circ Res.
2004; 94:802809.
Lancaster MK, Jones SA, Harrison SM, Boyett
MR. Intracellular Ca2+ and pacemaking within the
rabbit sinoatrial node: heterogeneity of role and
control. J Physiol. 2004; 556:481494.
16. Christoffels VM, Burch JB, Moorman AFM.
Architectural plan for the heart: early patterning
and delineation of the chambers and the nodes.
Trends Cardiovasc Med. 2004; 14:301307.
17. Maltsev VA, Vinogradova TM, Bogdanov KY,
Lakatta EG, Stern MD. Diastolic calcium release
controls the beating rate of rabbit sinoatrial node
cells: numerical modeling of the coupling
process. Biophys J. 2004; 86:25962605.
18. DiFrancesco D. The contribution of the
pacemaker current (if generation of spontaneous
activity in rabbit sino-atrial node myocytes. J
Physiol. 1991; 434:2340.
19. van Mierop LHS and Gessner IH. The
morphologic development of the sinoatrial node
in the mouse. Am J Cardiol. 1970; 25:204212.
20. Viragh Sz and Challice CE. The development of
the conduction system in the mouse embryo
heart. Dev Biol. 1980; 80:2845.
21. deJong F, Opthof T, Wilde AAM, Janse MJ,
Charles R, Lamers WH, Moorman AFM.
Persisting zones of slow impulse conduction in
developing chicken hearts. Circ Res. 1992;
71:240250.
22. Moorman AFM and Christoffels VM. Cardiac
chamber formation: development, genes, and
evolution. Physiol Rev. 2003; 83:12231267.
23. Mommersteeg MTM, Hoogaars WMH, Prall
OWJ, de Gier-de Vries C, Wiese C, Clout DEW,
Papaioannou VE, Brown NA, Harvey RP,
Moorman AFM, Christoffels VM. Molecular
pathway for the localized formation of the
sinoatrial node. Circ Res. 2007; 100:354362
24. Satin J, Itzhaki I, Rapoport S, Schroder EA, Izu
L, Arbel G, Beyar R, Balke CW, Schiller J,
Gepstein L. Calcium handling in human
embryonic stem cell derived cardiomyocytes.
Stem Cells. 2008; 26:19611972.
25. Konuri VK, Agnihotri G, Reddy BR. Current
advances and concepts of the embryological and
genetic basis of the developing human heart.
International Journal of Advanced Research
2014; 2: 431-435.
421
Vijaykumar et al.,
Int J Med Res Health Sci. 2015;4(2):417-421
You might also like
- Bamboos of India WebDocument354 pagesBamboos of India WebChristian Gauss100% (1)
- GR 10 Biology - Tissues Organs and System of Living ThingDocument28 pagesGR 10 Biology - Tissues Organs and System of Living Thingapi-301141568No ratings yet
- 100 Concepts of Developmental and Gross AnatomyDocument246 pages100 Concepts of Developmental and Gross AnatomyAbd Samad100% (1)
- Contractilidad Bombardini2005Document22 pagesContractilidad Bombardini2005Carlos Navarro alonsoNo ratings yet
- Jurnal 55Document1 pageJurnal 55rizalmantaNo ratings yet
- CalcioDocument31 pagesCalcioAlejandro Londoño AmarilesNo ratings yet
- Sarcoplasmic Reticulum 3Document6 pagesSarcoplasmic Reticulum 3Syaidah umiNo ratings yet
- UpToDate - Anti Arrhythmic Drugs MOADocument31 pagesUpToDate - Anti Arrhythmic Drugs MOA9821699080No ratings yet
- Calcium Sparks: Physiol RevDocument56 pagesCalcium Sparks: Physiol RevFisiologiaUFCGNo ratings yet
- Cardiac Pacing Learning Package 2013Document34 pagesCardiac Pacing Learning Package 2013Chetan Amberkar100% (1)
- 18sodium Calcium ExchangerDocument4 pages18sodium Calcium ExchangerZiedTrikiNo ratings yet
- Advanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Document28 pagesAdvanced 2physiology Assighnment On Automaticity & Rhythmici2ty (Autorecovered) (Autorecovered)Muhammad Mudassar ChudaryNo ratings yet
- Disorders of Heart RhythmsDocument202 pagesDisorders of Heart Rhythmspraveena.pv1234No ratings yet
- Cardiac Action Potential - WikipediaDocument67 pagesCardiac Action Potential - WikipediaMohd. Mudabbir-Ul- IslamNo ratings yet
- Principles of Cardiac ElectrophysiologyDocument98 pagesPrinciples of Cardiac ElectrophysiologyAbnet WondimuNo ratings yet
- Cardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateDocument32 pagesCardiac Excitability, Mechanisms of Arrhythmia, and Action of Antiarrhythmic Drugs - UpToDateClaudia AlessandraNo ratings yet
- Jphysiol 2009 181578Document2 pagesJphysiol 2009 181578ProtopaticoNo ratings yet
- Logy Assighnment On Automaticity Rhythmicity AutorecoveredDocument10 pagesLogy Assighnment On Automaticity Rhythmicity AutorecoveredM Asif RiazNo ratings yet
- Cardiac Action PotentialDocument5 pagesCardiac Action PotentialjayNo ratings yet
- Signaling in Muscle ContractionDocument15 pagesSignaling in Muscle ContractionMARTINEZ SANCHEZ JULIAN ELIASNo ratings yet
- CalcioDocument6 pagesCalcioArtur Franck Tenorio AronésNo ratings yet
- Physiology FAQ 2018Document185 pagesPhysiology FAQ 2018kirki pNo ratings yet
- Hypokalemia and Anesthetic Implications: Wong, Patrick Schafer, and John R. SchultzDocument23 pagesHypokalemia and Anesthetic Implications: Wong, Patrick Schafer, and John R. SchultzRidski D. MiruNo ratings yet
- Excitation PrintDocument3 pagesExcitation PrintZoRo4everNo ratings yet
- Heart PhysiolDocument84 pagesHeart PhysiolChintya Ayu ChampakaNo ratings yet
- L-Type Calcium ChannelDocument3 pagesL-Type Calcium ChannelNTA UGC-NETNo ratings yet
- Signaling in Muscle ContractionDocument14 pagesSignaling in Muscle ContractionJosué VelázquezNo ratings yet
- Nerves, Hormones and HomeostasisDocument8 pagesNerves, Hormones and Homeostasisumairas_1No ratings yet
- Oakes-Mitochondria Ca Immune SynapseDocument2 pagesOakes-Mitochondria Ca Immune Synapseiulia andreeaNo ratings yet
- Cytosolic Calcium Homeostasis in Neurons - Control Systems, Modulation by Reactive Oxygen and Nitrogen Species, and Space and Time FluctuationsDocument52 pagesCytosolic Calcium Homeostasis in Neurons - Control Systems, Modulation by Reactive Oxygen and Nitrogen Species, and Space and Time Fluctuationskj185No ratings yet
- Channelopathies: Ion Channel Defects Linked To Heritable Clinical DisordersDocument12 pagesChannelopathies: Ion Channel Defects Linked To Heritable Clinical Disorderscote_cote6951No ratings yet
- Llinas PNASDocument6 pagesLlinas PNASjhrubioNo ratings yet
- Molecular Basis of Cardiac Action Potential RepolarizationDocument6 pagesMolecular Basis of Cardiac Action Potential RepolarizationJuan Jose Valladares SanchezNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologySteve FerisNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologyTamjid Khan100% (2)
- Arrythmia by AbnetDocument509 pagesArrythmia by AbnetAbnet WondimuNo ratings yet
- Calcium in BiologyDocument8 pagesCalcium in BiologyomNo ratings yet
- Review Article Crosstalk Between Calcium and ROS in Pathophysiological ConditionsDocument19 pagesReview Article Crosstalk Between Calcium and ROS in Pathophysiological ConditionsbetiNo ratings yet
- Action PotentialDocument2 pagesAction PotentialMian Talha KamalNo ratings yet
- Cardiac Muscle Physiology: P. Szedlak, D.S. Steele and P.M. HopkinsDocument8 pagesCardiac Muscle Physiology: P. Szedlak, D.S. Steele and P.M. Hopkinsmastmulla921No ratings yet
- Tissue To Organisms/Physiology & PharmacologyDocument6 pagesTissue To Organisms/Physiology & PharmacologyStudent LifeNo ratings yet
- Praktikum Faal-Blok14-3BDocument18 pagesPraktikum Faal-Blok14-3BKelompok 5A Semester 4No ratings yet
- Exercise-Induced Rhabdomyolysis Mechanisms and PreventionDocument11 pagesExercise-Induced Rhabdomyolysis Mechanisms and PreventionMarlene PgNo ratings yet
- Cellular Biology of EpileptogenesisDocument10 pagesCellular Biology of EpileptogenesisandreacamachogonNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsleandronascciNo ratings yet
- QSAR Studies On Calcium Channel Blockers: Satya P. GuptaDocument39 pagesQSAR Studies On Calcium Channel Blockers: Satya P. GuptaOana FloreanNo ratings yet
- UNIT 9 NotesDocument5 pagesUNIT 9 NotesMariaNo ratings yet
- The Journal of Physiology - 2008 - Berridge - Smooth Muscle Cell Calcium Activation MechanismsDocument15 pagesThe Journal of Physiology - 2008 - Berridge - Smooth Muscle Cell Calcium Activation MechanismsNoumanNo ratings yet
- Human Physiology Assignment 01Document9 pagesHuman Physiology Assignment 01Evaristo MK VincentNo ratings yet
- The Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesDocument30 pagesThe Journal of Physiology - 1967 - Katz - A Study of Synaptic Transmission in The Absence of Nerve ImpulsesKhel VincentNo ratings yet
- Pi Is 0022282817303115Document11 pagesPi Is 0022282817303115EdgarNo ratings yet
- Neurochem FinalDocument20 pagesNeurochem Finalelifulku.a1No ratings yet
- 1.bomba de Na - KDocument22 pages1.bomba de Na - KJosé DiegoNo ratings yet
- Potassium Channels ThesisDocument7 pagesPotassium Channels Thesisbk156rhq100% (2)
- Computer SimulationDocument29 pagesComputer SimulationmrtiborhornyikNo ratings yet
- Skeletal Muscle Physiology PDFDocument6 pagesSkeletal Muscle Physiology PDFAstri Ggamjong Xiao LuNo ratings yet
- 2 02-Electrocardiography PDFDocument17 pages2 02-Electrocardiography PDFMiguel DomingoNo ratings yet
- Cardiac PDFDocument49 pagesCardiac PDFViswani pillapalyamNo ratings yet
- Cardiac Action PotentialDocument41 pagesCardiac Action PotentialRadijska Postaja KoprivnicaNo ratings yet
- Advanced Cell BiologyDocument14 pagesAdvanced Cell BiologyUmair AneesNo ratings yet
- Learning Objectives: Chapter 4: Electrophysiology of The HeartDocument28 pagesLearning Objectives: Chapter 4: Electrophysiology of The HeartLaura PucheNo ratings yet
- DISCUDocument23 pagesDISCUpvsvrNo ratings yet
- EKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXFrom EverandEKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXRating: 5 out of 5 stars5/5 (2)
- Ijmrhs Vol 4 Issue 2Document219 pagesIjmrhs Vol 4 Issue 2editorijmrhsNo ratings yet
- Ijmrhs Vol 4 Issue 4Document193 pagesIjmrhs Vol 4 Issue 4editorijmrhs0% (1)
- Ijmrhs Vol 4 Issue 3Document263 pagesIjmrhs Vol 4 Issue 3editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 4Document294 pagesIjmrhs Vol 3 Issue 4editorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 4Document321 pagesIjmrhs Vol 2 Issue 4editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 2Document281 pagesIjmrhs Vol 3 Issue 2editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 1Document228 pagesIjmrhs Vol 3 Issue 1editorijmrhsNo ratings yet
- Ijmrhs Vol 3 Issue 3Document271 pagesIjmrhs Vol 3 Issue 3editorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 3Document399 pagesIjmrhs Vol 2 Issue 3editorijmrhs100% (1)
- Ijmrhs Vol 1 Issue 1Document257 pagesIjmrhs Vol 1 Issue 1editorijmrhsNo ratings yet
- 48 MakrandDocument2 pages48 MakrandeditorijmrhsNo ratings yet
- Ijmrhs Vol 2 Issue 1Document110 pagesIjmrhs Vol 2 Issue 1editorijmrhs100% (1)
- Ijmrhs Vol 2 Issue 2Document197 pagesIjmrhs Vol 2 Issue 2editorijmrhsNo ratings yet
- 35krishnasamy EtalDocument1 page35krishnasamy EtaleditorijmrhsNo ratings yet
- 47serban Turliuc EtalDocument4 pages47serban Turliuc EtaleditorijmrhsNo ratings yet
- Recurrent Cornual Ectopic Pregnancy - A Case Report: Article InfoDocument2 pagesRecurrent Cornual Ectopic Pregnancy - A Case Report: Article InfoeditorijmrhsNo ratings yet
- 45mohit EtalDocument4 pages45mohit EtaleditorijmrhsNo ratings yet
- Pernicious Anemia in Young: A Case Report With Review of LiteratureDocument5 pagesPernicious Anemia in Young: A Case Report With Review of LiteratureeditorijmrhsNo ratings yet
- Williams-Campbell Syndrome-A Rare Entity of Congenital Bronchiectasis: A Case Report in AdultDocument3 pagesWilliams-Campbell Syndrome-A Rare Entity of Congenital Bronchiectasis: A Case Report in AdulteditorijmrhsNo ratings yet
- 36rashmipal EtalDocument6 pages36rashmipal EtaleditorijmrhsNo ratings yet
- 37poflee EtalDocument3 pages37poflee EtaleditorijmrhsNo ratings yet
- 38vaishnavi EtalDocument3 pages38vaishnavi EtaleditorijmrhsNo ratings yet
- 41anurag EtalDocument2 pages41anurag EtaleditorijmrhsNo ratings yet
- 40vedant EtalDocument4 pages40vedant EtaleditorijmrhsNo ratings yet
- 31tushar EtalDocument4 pages31tushar EtaleditorijmrhsNo ratings yet
- 34tupe EtalDocument5 pages34tupe EtaleditorijmrhsNo ratings yet
- 33 Prabu RamDocument5 pages33 Prabu RameditorijmrhsNo ratings yet
- 28nnadi EtalDocument4 pages28nnadi EtaleditorijmrhsNo ratings yet
- Introduction (Recovered)Document37 pagesIntroduction (Recovered)muhamedshuaibbhaiNo ratings yet
- Anatomy of The Nervous SystemDocument85 pagesAnatomy of The Nervous SystemjoelNo ratings yet
- Johnson Et Al.2006. Felidae Phylogeny PDFDocument6 pagesJohnson Et Al.2006. Felidae Phylogeny PDFNydia JGNo ratings yet
- Bamboo Species Source List No. 36 2017-2018Document56 pagesBamboo Species Source List No. 36 2017-2018André MichaelNo ratings yet
- Nettle FibersDocument7 pagesNettle Fibersawais123142541254125No ratings yet
- Arthroscopy of The Temporomandibular JointDocument44 pagesArthroscopy of The Temporomandibular JointDencyclopediaNo ratings yet
- Classification of OrganismDocument35 pagesClassification of OrganismJnll JstnNo ratings yet
- SIPDocument20 pagesSIPLuiiiNo ratings yet
- Grade 11 Q2 Module 6 Week 6Document11 pagesGrade 11 Q2 Module 6 Week 6Shiela MaeNo ratings yet
- Grassland Landscape ManagementDocument22 pagesGrassland Landscape ManagementAr Deepika RajaNo ratings yet
- The Fundamental Unit of LifeDocument16 pagesThe Fundamental Unit of LifeSriram RNo ratings yet
- Type-I Collagen Amino Acid Glycine Collagen Triple HelixDocument8 pagesType-I Collagen Amino Acid Glycine Collagen Triple Helixkyle swayNo ratings yet
- Hill Stream AdaptationDocument5 pagesHill Stream AdaptationAdarsh SenNo ratings yet
- Anxiety Lesson WorksheetDocument4 pagesAnxiety Lesson WorksheetAmisha VastaniNo ratings yet
- Recommended Procedures For Sample Collection Preservation and Shipping-D.senneDocument48 pagesRecommended Procedures For Sample Collection Preservation and Shipping-D.senneAndersonNo ratings yet
- Skin Conditions Worksheet 2017Document3 pagesSkin Conditions Worksheet 2017Tamia WatsonNo ratings yet
- AnthDocument18 pagesAnthAnurag ShuklaNo ratings yet
- Evolution of AnuraDocument18 pagesEvolution of AnuraSan Nguyễn ViếtNo ratings yet
- Nat 5 Biology SQ App 2019Document48 pagesNat 5 Biology SQ App 2019muntazNo ratings yet
- New English Sba 2018Document15 pagesNew English Sba 2018ashawni patterson88% (8)
- 2024 Msyi Final ProgramDocument12 pages2024 Msyi Final Programapi-654587216No ratings yet
- Result Oriented Management (ROM) - 2Document7 pagesResult Oriented Management (ROM) - 2baguma100% (1)
- Biology and Physiology of Saline PlantsDocument32 pagesBiology and Physiology of Saline PlantsNguyễn Lê Quốc ThiNo ratings yet
- Scientific Aspects in Ramcharit ManasDocument52 pagesScientific Aspects in Ramcharit ManasRam Kumar Kuril100% (9)
- 16625.Hsl4207 Microbiology (Parasitology & Mycology) CH 13Document4 pages16625.Hsl4207 Microbiology (Parasitology & Mycology) CH 13Tarun AroraNo ratings yet
- Assessment Diagnosis Scientific Reason Planning Intervention Rational E EvaluationDocument2 pagesAssessment Diagnosis Scientific Reason Planning Intervention Rational E EvaluationCamille VirayNo ratings yet
- Supplement Guide SleepDocument39 pagesSupplement Guide SleepinfoNo ratings yet