
Velez 2013
Velez 2013
You might also like
- Unlocking The Mystery of Life ScriptDocument47 pagesUnlocking The Mystery of Life ScriptmimikashynninNo ratings yet
- Activated Sludge Troubleshooting ChartDocument1 pageActivated Sludge Troubleshooting ChartIslandHighlander100% (2)
- Scalar Wave Energy As Weapon: June 2019Document54 pagesScalar Wave Energy As Weapon: June 2019allhigorNo ratings yet
- The Long Time Dynamics of Two HydrodynamicallyDocument33 pagesThe Long Time Dynamics of Two HydrodynamicallyKaterina PapadopoulouNo ratings yet
- Fung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHDocument6 pagesFung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHcarciclaroNo ratings yet
- Jun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaDocument11 pagesJun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaGretymjNo ratings yet
- Life at Low Reynolds NumberDocument9 pagesLife at Low Reynolds Numberapi-238456622No ratings yet
- 1 s2.0 S2211379717312421 MainDocument8 pages1 s2.0 S2211379717312421 MainAdil IhsanNo ratings yet
- Active Oscillations in Microscale Navigation: Kirsty Y. WanDocument14 pagesActive Oscillations in Microscale Navigation: Kirsty Y. WanAnonymous KoAJZdB8No ratings yet
- Fluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesDocument19 pagesFluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesSantiago ToroNo ratings yet
- Controlled Locomotion of A Droplet Propelled by An Encapsulated SquirmerDocument9 pagesControlled Locomotion of A Droplet Propelled by An Encapsulated SquirmerArindam BasakNo ratings yet
- Comparison of SwimmerDocument11 pagesComparison of SwimmerTreeNo ratings yet
- Swimming PDFDocument14 pagesSwimming PDF917435001No ratings yet
- Microscopic Artificial Swimmers - Rémi Dreyfus Et AlDocument5 pagesMicroscopic Artificial Swimmers - Rémi Dreyfus Et Alalfredc20No ratings yet
- On The Best Design For Undulatory Swimming: Christophe EloyDocument41 pagesOn The Best Design For Undulatory Swimming: Christophe EloyZuo CuiNo ratings yet
- Osmosis Through A Semi-Permeable Membrane: A Consistent Approach To InteractionsDocument36 pagesOsmosis Through A Semi-Permeable Membrane: A Consistent Approach To InteractionsBuxNo ratings yet
- Chung2019 Article AMinireviewOnInertialMicrofluiDocument11 pagesChung2019 Article AMinireviewOnInertialMicrofluiKottapalli Sundar mahesh (RA1811034010048)No ratings yet
- A Minireview On Inertial Microfluidics Fundamentals: Inertial Particle Focusing and Secondary FlowDocument2 pagesA Minireview On Inertial Microfluidics Fundamentals: Inertial Particle Focusing and Secondary FlowRaul Alejandro Ramirez GomezNo ratings yet
- Journal FreeDocument36 pagesJournal FreeRiccardo Spinoza SilvaNo ratings yet
- A Review On Gyrotactic Swimmers in Turbulent Flows: Acta Mechanica SinicaDocument14 pagesA Review On Gyrotactic Swimmers in Turbulent Flows: Acta Mechanica SinicaBadar E AlamNo ratings yet
- Hydrostatic Pressure and Interfacial Tension Induce Mode Instability in Wave Propagation Along A Liquid-Filled MicrotubuleDocument11 pagesHydrostatic Pressure and Interfacial Tension Induce Mode Instability in Wave Propagation Along A Liquid-Filled MicrotubuleDejanNo ratings yet
- Wadhwa Et Al-2021-Nature Reviews Microbiology-1Document13 pagesWadhwa Et Al-2021-Nature Reviews Microbiology-1pridNo ratings yet
- Pulsing Corals: A Story of Scale and Mixing: Original ArticleDocument14 pagesPulsing Corals: A Story of Scale and Mixing: Original ArticleWyatt GrayNo ratings yet
- New Journal of Physics: Hydrodynamic Theory For Multi-Component Active Polar GelsDocument17 pagesNew Journal of Physics: Hydrodynamic Theory For Multi-Component Active Polar GelsAbdelkebir Es-salmiNo ratings yet
- Weak Synchronization and Large-Scale Collective OscillationDocument5 pagesWeak Synchronization and Large-Scale Collective OscillationTrung PhanNo ratings yet
- Ves 5Document16 pagesVes 5Khushii NaamdeoNo ratings yet
- DePietro 2015Document8 pagesDePietro 2015pedroNo ratings yet
- Capillary-Based Static Self-Assembly in Higher Organisms: ReferencesDocument11 pagesCapillary-Based Static Self-Assembly in Higher Organisms: ReferencesTalal Al-HousseinyNo ratings yet
- TPH 1Document19 pagesTPH 1ali hadiNo ratings yet
- Contact Angle, Wettability and Adhesion (Vol 3) - K. Mittal (VSP, 2003) WWDocument513 pagesContact Angle, Wettability and Adhesion (Vol 3) - K. Mittal (VSP, 2003) WWkarlGG100% (2)
- The Hydrodynamics of A Run-and-Tumble Bacterium Propelled by Polymorphic Helical FlagellaDocument6 pagesThe Hydrodynamics of A Run-and-Tumble Bacterium Propelled by Polymorphic Helical FlagellaFernandoNo ratings yet
- 1 s2.0 S0045793012003027 MainDocument17 pages1 s2.0 S0045793012003027 MainZaka MuhammadNo ratings yet
- Multi-Particle Collision Dynamics: Flow Around A Circular and A Square CylinderDocument9 pagesMulti-Particle Collision Dynamics: Flow Around A Circular and A Square CylinderEwerton AvlisNo ratings yet
- Microswimming in Viscoelastic FluidsDocument16 pagesMicroswimming in Viscoelastic FluidsMuhammad MusharafNo ratings yet
- Sachin Goyal, N. C. Perkins and Christopher L. Lee - Nonlinear Dynamic Intertwining of Rods With Self-ContactDocument35 pagesSachin Goyal, N. C. Perkins and Christopher L. Lee - Nonlinear Dynamic Intertwining of Rods With Self-ContactLokosooNo ratings yet
- Micro Fluidics PaperDocument12 pagesMicro Fluidics PaperBlake SteinesNo ratings yet
- Using Lagrangian Coherent Structures To Analyze Fluid Mixing by CiliaDocument9 pagesUsing Lagrangian Coherent Structures To Analyze Fluid Mixing by Ciliazsong953No ratings yet
- Singh, Kinematic WaveDocument50 pagesSingh, Kinematic Wavearanguiz2012No ratings yet
- Interplay Between Mechanosensitive Adhesions and MDocument13 pagesInterplay Between Mechanosensitive Adhesions and MPinaki NayakNo ratings yet
- ScalarWaveDrivenEnergyApplications Chapter6Document49 pagesScalarWaveDrivenEnergyApplications Chapter6lwNo ratings yet
- Maximum Propulsive Swimming WakesDocument10 pagesMaximum Propulsive Swimming WakesCezar BolzanNo ratings yet
- Cell MigrationDocument9 pagesCell MigrationGordon YapNo ratings yet
- Keshet Swarm Modeling 2001Document7 pagesKeshet Swarm Modeling 2001Aakash SarkarNo ratings yet
- Chaotic Dynamics and Oxygen Transport in Thin Films of Aerotactic BacteriaDocument8 pagesChaotic Dynamics and Oxygen Transport in Thin Films of Aerotactic BacteriaJamie SamuelNo ratings yet
- Scalar WavesDocument49 pagesScalar WavesEduardoNo ratings yet
- 2013-POF-nonlinear Oscillations Following The Rayleigh Collapse of A Gas Bubble in A Linear Viscoelastic MediumDocument23 pages2013-POF-nonlinear Oscillations Following The Rayleigh Collapse of A Gas Bubble in A Linear Viscoelastic MediumzyxNo ratings yet
- EnergyRateDensity II Galley 2011Document20 pagesEnergyRateDensity II Galley 2011Tyler PooleNo ratings yet
- 2014 Discontinuity-Induced Bifurcation Cascades in Flows and Maps - DankowiczDocument16 pages2014 Discontinuity-Induced Bifurcation Cascades in Flows and Maps - DankowiczFabian GarciaNo ratings yet
- Huashu Dou Origin of Turbulence Energy Gradient TheoryDocument503 pagesHuashu Dou Origin of Turbulence Energy Gradient TheoryStrahinja DonicNo ratings yet
- Swimming by Reciprocal Motion at Low Reynolds Number: ArticleDocument8 pagesSwimming by Reciprocal Motion at Low Reynolds Number: Articlejuan felipeNo ratings yet
- CRAS WhaleDocument13 pagesCRAS WhaleRNo ratings yet
- 694 FullDocument9 pages694 FullRika Rahma PutriNo ratings yet
- Complex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesDocument7 pagesComplex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesGretymjNo ratings yet
- Scalar Wave Energy As Weapon: June 2019Document54 pagesScalar Wave Energy As Weapon: June 2019allhigorNo ratings yet
- Rough DraftDocument18 pagesRough DraftVikram ChavanNo ratings yet
- Biomimetics: The Ground Effect in Anguilliform SwimmingDocument13 pagesBiomimetics: The Ground Effect in Anguilliform SwimmingimanborazjaniNo ratings yet
- Corresponding Author: J.G. Cervantes: Jgonzalo@servidot - Unam.mxDocument20 pagesCorresponding Author: J.G. Cervantes: Jgonzalo@servidot - Unam.mxJaime Cervantes de GortariNo ratings yet
- FD Att TechtalkDocument12 pagesFD Att TechtalkpraneethNo ratings yet
- Muller Larval Flow JEB08Document11 pagesMuller Larval Flow JEB08Mohamed Al-Sayed Al-GendyNo ratings yet
- Robotic Models For Studying Undulatory Locomotion in Fishes: PaperDocument15 pagesRobotic Models For Studying Undulatory Locomotion in Fishes: PaperLucaNo ratings yet
- Disipación ViscosaDocument5 pagesDisipación ViscosajucasegaNo ratings yet
- High Values of CLDocument12 pagesHigh Values of CLjucasegaNo ratings yet
- Char LatexDocument1 pageChar LatexjucasegaNo ratings yet
- Rotational DynamicsDocument6 pagesRotational DynamicsjucasegaNo ratings yet
- Competition and Interaction of Polydisperse Bubbles in Polymer FoamsDocument12 pagesCompetition and Interaction of Polydisperse Bubbles in Polymer FoamsjucasegaNo ratings yet
- Edelwise A. Espada Cell ViewingDocument4 pagesEdelwise A. Espada Cell ViewingAshley Villarez PlatonNo ratings yet
- Differences Between Prokaryotic & Eukaryotic Cells PresentationDocument24 pagesDifferences Between Prokaryotic & Eukaryotic Cells PresentationGeoffrey100% (1)
- Basic Bacteriology: MunawirDocument50 pagesBasic Bacteriology: MunawirindahirmawatiNo ratings yet
- JetDocument14 pagesJetAnnie Lyn VillamorNo ratings yet
- Lynn Margulis, Karlene Schwartz, Michael Dolan - DiversityDocument252 pagesLynn Margulis, Karlene Schwartz, Michael Dolan - DiversityJavier Raksha100% (1)
- Cilia and Flagella: Structure and Function in EukaryotesDocument10 pagesCilia and Flagella: Structure and Function in EukaryotesEr Purushottam PalNo ratings yet
- Micropara TransesDocument16 pagesMicropara TransesethylnaaadesNo ratings yet
- Eukaryote - WikipediaDocument19 pagesEukaryote - WikipediaEllysa Mae Capinig RiedlerNo ratings yet
- The Flagellated ProtozoansDocument27 pagesThe Flagellated ProtozoansJolaine ValloNo ratings yet
- Bacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaDocument4 pagesBacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaTLCRNo ratings yet
- Post Lab Report - Exercise 1 - Morphology and Motility of MicroorganismsDocument23 pagesPost Lab Report - Exercise 1 - Morphology and Motility of MicroorganismsDara KwonNo ratings yet
- Structure of MicrobesDocument118 pagesStructure of MicrobesDeana Namirembe100% (6)
- Microbiology: PROKARYOTIC & EUKARYOTIC CELLSDocument10 pagesMicrobiology: PROKARYOTIC & EUKARYOTIC CELLSEmily RaeNo ratings yet
- Bacterial Disease AnalysisDocument63 pagesBacterial Disease AnalysisAshutoshkumar jhaNo ratings yet
- SHS - Biology 1 - Q1 - Module 3 - Reviewed - EditedDocument30 pagesSHS - Biology 1 - Q1 - Module 3 - Reviewed - EditedLiberty Salazar100% (2)
- BIO 275 - Chapter 3 and 4 QuestionsDocument3 pagesBIO 275 - Chapter 3 and 4 QuestionsLindsey StilleyNo ratings yet
- F. Y. B. Sc. (Microbiology) Paper-I Question Bank PDFDocument16 pagesF. Y. B. Sc. (Microbiology) Paper-I Question Bank PDFShipra ChakrabortyNo ratings yet
- Altas de Dasicladaceas P 1-50Document50 pagesAltas de Dasicladaceas P 1-508811791No ratings yet
- Comparing Extracellular Secretion Mechanisms in E. Coli Strains BL21 (DE3) and Nissle 1917Document10 pagesComparing Extracellular Secretion Mechanisms in E. Coli Strains BL21 (DE3) and Nissle 1917vikramreddy041302No ratings yet
- Soal ToeflDocument11 pagesSoal ToeflNurul A'la RojasNo ratings yet
- Chapter 4: ProtistaDocument8 pagesChapter 4: ProtistaNawal Che IsmailNo ratings yet
- Bio 1 PDFDocument13 pagesBio 1 PDFjayan sivaNo ratings yet
- What Are Cell OrganellesDocument19 pagesWhat Are Cell OrganellesMaraon CharitaNo ratings yet
- Important Questions For CBSE Class 11 Biology Chapter 8Document15 pagesImportant Questions For CBSE Class 11 Biology Chapter 8lavanya rajaNo ratings yet
- Anatomy of Bacteria: Morphology Structure FunctionDocument49 pagesAnatomy of Bacteria: Morphology Structure FunctionBlanche AltheaNo ratings yet
- Cell StructersDocument13 pagesCell StructersSumaiya JabinNo ratings yet
- Topic: Bacteria: EtymologyDocument28 pagesTopic: Bacteria: EtymologyVINDHYA SHANKERNo ratings yet
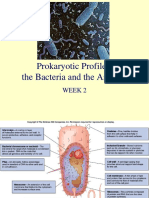
- Prokaryotic Profiles: The Bacteria and The Archaea: Week 2Document44 pagesProkaryotic Profiles: The Bacteria and The Archaea: Week 2Irwan M. IskoberNo ratings yet




























































































Download as pdf or txt
You might also like
- Unlocking The Mystery of Life ScriptDocument47 pagesUnlocking The Mystery of Life ScriptmimikashynninNo ratings yet
- Activated Sludge Troubleshooting ChartDocument1 pageActivated Sludge Troubleshooting ChartIslandHighlander100% (2)
- Scalar Wave Energy As Weapon: June 2019Document54 pagesScalar Wave Energy As Weapon: June 2019allhigorNo ratings yet
- The Long Time Dynamics of Two HydrodynamicallyDocument33 pagesThe Long Time Dynamics of Two HydrodynamicallyKaterina PapadopoulouNo ratings yet
- Fung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHDocument6 pagesFung Et Al 2022 Swimming Feeding and Inversion of Multicellular CHcarciclaroNo ratings yet
- Jun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaDocument11 pagesJun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaGretymjNo ratings yet
- Life at Low Reynolds NumberDocument9 pagesLife at Low Reynolds Numberapi-238456622No ratings yet
- 1 s2.0 S2211379717312421 MainDocument8 pages1 s2.0 S2211379717312421 MainAdil IhsanNo ratings yet
- Active Oscillations in Microscale Navigation: Kirsty Y. WanDocument14 pagesActive Oscillations in Microscale Navigation: Kirsty Y. WanAnonymous KoAJZdB8No ratings yet
- Fluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesDocument19 pagesFluid Flows and Forces in Development Functions, Features and Biophysical PrinciplesSantiago ToroNo ratings yet
- Controlled Locomotion of A Droplet Propelled by An Encapsulated SquirmerDocument9 pagesControlled Locomotion of A Droplet Propelled by An Encapsulated SquirmerArindam BasakNo ratings yet
- Comparison of SwimmerDocument11 pagesComparison of SwimmerTreeNo ratings yet
- Swimming PDFDocument14 pagesSwimming PDF917435001No ratings yet
- Microscopic Artificial Swimmers - Rémi Dreyfus Et AlDocument5 pagesMicroscopic Artificial Swimmers - Rémi Dreyfus Et Alalfredc20No ratings yet
- On The Best Design For Undulatory Swimming: Christophe EloyDocument41 pagesOn The Best Design For Undulatory Swimming: Christophe EloyZuo CuiNo ratings yet
- Osmosis Through A Semi-Permeable Membrane: A Consistent Approach To InteractionsDocument36 pagesOsmosis Through A Semi-Permeable Membrane: A Consistent Approach To InteractionsBuxNo ratings yet
- Chung2019 Article AMinireviewOnInertialMicrofluiDocument11 pagesChung2019 Article AMinireviewOnInertialMicrofluiKottapalli Sundar mahesh (RA1811034010048)No ratings yet
- A Minireview On Inertial Microfluidics Fundamentals: Inertial Particle Focusing and Secondary FlowDocument2 pagesA Minireview On Inertial Microfluidics Fundamentals: Inertial Particle Focusing and Secondary FlowRaul Alejandro Ramirez GomezNo ratings yet
- Journal FreeDocument36 pagesJournal FreeRiccardo Spinoza SilvaNo ratings yet
- A Review On Gyrotactic Swimmers in Turbulent Flows: Acta Mechanica SinicaDocument14 pagesA Review On Gyrotactic Swimmers in Turbulent Flows: Acta Mechanica SinicaBadar E AlamNo ratings yet
- Hydrostatic Pressure and Interfacial Tension Induce Mode Instability in Wave Propagation Along A Liquid-Filled MicrotubuleDocument11 pagesHydrostatic Pressure and Interfacial Tension Induce Mode Instability in Wave Propagation Along A Liquid-Filled MicrotubuleDejanNo ratings yet
- Wadhwa Et Al-2021-Nature Reviews Microbiology-1Document13 pagesWadhwa Et Al-2021-Nature Reviews Microbiology-1pridNo ratings yet
- Pulsing Corals: A Story of Scale and Mixing: Original ArticleDocument14 pagesPulsing Corals: A Story of Scale and Mixing: Original ArticleWyatt GrayNo ratings yet
- New Journal of Physics: Hydrodynamic Theory For Multi-Component Active Polar GelsDocument17 pagesNew Journal of Physics: Hydrodynamic Theory For Multi-Component Active Polar GelsAbdelkebir Es-salmiNo ratings yet
- Weak Synchronization and Large-Scale Collective OscillationDocument5 pagesWeak Synchronization and Large-Scale Collective OscillationTrung PhanNo ratings yet
- Ves 5Document16 pagesVes 5Khushii NaamdeoNo ratings yet
- DePietro 2015Document8 pagesDePietro 2015pedroNo ratings yet
- Capillary-Based Static Self-Assembly in Higher Organisms: ReferencesDocument11 pagesCapillary-Based Static Self-Assembly in Higher Organisms: ReferencesTalal Al-HousseinyNo ratings yet
- TPH 1Document19 pagesTPH 1ali hadiNo ratings yet
- Contact Angle, Wettability and Adhesion (Vol 3) - K. Mittal (VSP, 2003) WWDocument513 pagesContact Angle, Wettability and Adhesion (Vol 3) - K. Mittal (VSP, 2003) WWkarlGG100% (2)
- The Hydrodynamics of A Run-and-Tumble Bacterium Propelled by Polymorphic Helical FlagellaDocument6 pagesThe Hydrodynamics of A Run-and-Tumble Bacterium Propelled by Polymorphic Helical FlagellaFernandoNo ratings yet
- 1 s2.0 S0045793012003027 MainDocument17 pages1 s2.0 S0045793012003027 MainZaka MuhammadNo ratings yet
- Multi-Particle Collision Dynamics: Flow Around A Circular and A Square CylinderDocument9 pagesMulti-Particle Collision Dynamics: Flow Around A Circular and A Square CylinderEwerton AvlisNo ratings yet
- Microswimming in Viscoelastic FluidsDocument16 pagesMicroswimming in Viscoelastic FluidsMuhammad MusharafNo ratings yet
- Sachin Goyal, N. C. Perkins and Christopher L. Lee - Nonlinear Dynamic Intertwining of Rods With Self-ContactDocument35 pagesSachin Goyal, N. C. Perkins and Christopher L. Lee - Nonlinear Dynamic Intertwining of Rods With Self-ContactLokosooNo ratings yet
- Micro Fluidics PaperDocument12 pagesMicro Fluidics PaperBlake SteinesNo ratings yet
- Using Lagrangian Coherent Structures To Analyze Fluid Mixing by CiliaDocument9 pagesUsing Lagrangian Coherent Structures To Analyze Fluid Mixing by Ciliazsong953No ratings yet
- Singh, Kinematic WaveDocument50 pagesSingh, Kinematic Wavearanguiz2012No ratings yet
- Interplay Between Mechanosensitive Adhesions and MDocument13 pagesInterplay Between Mechanosensitive Adhesions and MPinaki NayakNo ratings yet
- ScalarWaveDrivenEnergyApplications Chapter6Document49 pagesScalarWaveDrivenEnergyApplications Chapter6lwNo ratings yet
- Maximum Propulsive Swimming WakesDocument10 pagesMaximum Propulsive Swimming WakesCezar BolzanNo ratings yet
- Cell MigrationDocument9 pagesCell MigrationGordon YapNo ratings yet
- Keshet Swarm Modeling 2001Document7 pagesKeshet Swarm Modeling 2001Aakash SarkarNo ratings yet
- Chaotic Dynamics and Oxygen Transport in Thin Films of Aerotactic BacteriaDocument8 pagesChaotic Dynamics and Oxygen Transport in Thin Films of Aerotactic BacteriaJamie SamuelNo ratings yet
- Scalar WavesDocument49 pagesScalar WavesEduardoNo ratings yet
- 2013-POF-nonlinear Oscillations Following The Rayleigh Collapse of A Gas Bubble in A Linear Viscoelastic MediumDocument23 pages2013-POF-nonlinear Oscillations Following The Rayleigh Collapse of A Gas Bubble in A Linear Viscoelastic MediumzyxNo ratings yet
- EnergyRateDensity II Galley 2011Document20 pagesEnergyRateDensity II Galley 2011Tyler PooleNo ratings yet
- 2014 Discontinuity-Induced Bifurcation Cascades in Flows and Maps - DankowiczDocument16 pages2014 Discontinuity-Induced Bifurcation Cascades in Flows and Maps - DankowiczFabian GarciaNo ratings yet
- Huashu Dou Origin of Turbulence Energy Gradient TheoryDocument503 pagesHuashu Dou Origin of Turbulence Energy Gradient TheoryStrahinja DonicNo ratings yet
- Swimming by Reciprocal Motion at Low Reynolds Number: ArticleDocument8 pagesSwimming by Reciprocal Motion at Low Reynolds Number: Articlejuan felipeNo ratings yet
- CRAS WhaleDocument13 pagesCRAS WhaleRNo ratings yet
- 694 FullDocument9 pages694 FullRika Rahma PutriNo ratings yet
- Complex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesDocument7 pagesComplex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesGretymjNo ratings yet
- Scalar Wave Energy As Weapon: June 2019Document54 pagesScalar Wave Energy As Weapon: June 2019allhigorNo ratings yet
- Rough DraftDocument18 pagesRough DraftVikram ChavanNo ratings yet
- Biomimetics: The Ground Effect in Anguilliform SwimmingDocument13 pagesBiomimetics: The Ground Effect in Anguilliform SwimmingimanborazjaniNo ratings yet
- Corresponding Author: J.G. Cervantes: Jgonzalo@servidot - Unam.mxDocument20 pagesCorresponding Author: J.G. Cervantes: Jgonzalo@servidot - Unam.mxJaime Cervantes de GortariNo ratings yet
- FD Att TechtalkDocument12 pagesFD Att TechtalkpraneethNo ratings yet
- Muller Larval Flow JEB08Document11 pagesMuller Larval Flow JEB08Mohamed Al-Sayed Al-GendyNo ratings yet
- Robotic Models For Studying Undulatory Locomotion in Fishes: PaperDocument15 pagesRobotic Models For Studying Undulatory Locomotion in Fishes: PaperLucaNo ratings yet
- Disipación ViscosaDocument5 pagesDisipación ViscosajucasegaNo ratings yet
- High Values of CLDocument12 pagesHigh Values of CLjucasegaNo ratings yet
- Char LatexDocument1 pageChar LatexjucasegaNo ratings yet
- Rotational DynamicsDocument6 pagesRotational DynamicsjucasegaNo ratings yet
- Competition and Interaction of Polydisperse Bubbles in Polymer FoamsDocument12 pagesCompetition and Interaction of Polydisperse Bubbles in Polymer FoamsjucasegaNo ratings yet
- Edelwise A. Espada Cell ViewingDocument4 pagesEdelwise A. Espada Cell ViewingAshley Villarez PlatonNo ratings yet
- Differences Between Prokaryotic & Eukaryotic Cells PresentationDocument24 pagesDifferences Between Prokaryotic & Eukaryotic Cells PresentationGeoffrey100% (1)
- Basic Bacteriology: MunawirDocument50 pagesBasic Bacteriology: MunawirindahirmawatiNo ratings yet
- JetDocument14 pagesJetAnnie Lyn VillamorNo ratings yet
- Lynn Margulis, Karlene Schwartz, Michael Dolan - DiversityDocument252 pagesLynn Margulis, Karlene Schwartz, Michael Dolan - DiversityJavier Raksha100% (1)
- Cilia and Flagella: Structure and Function in EukaryotesDocument10 pagesCilia and Flagella: Structure and Function in EukaryotesEr Purushottam PalNo ratings yet
- Micropara TransesDocument16 pagesMicropara TransesethylnaaadesNo ratings yet
- Eukaryote - WikipediaDocument19 pagesEukaryote - WikipediaEllysa Mae Capinig RiedlerNo ratings yet
- The Flagellated ProtozoansDocument27 pagesThe Flagellated ProtozoansJolaine ValloNo ratings yet
- Bacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaDocument4 pagesBacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaTLCRNo ratings yet
- Post Lab Report - Exercise 1 - Morphology and Motility of MicroorganismsDocument23 pagesPost Lab Report - Exercise 1 - Morphology and Motility of MicroorganismsDara KwonNo ratings yet
- Structure of MicrobesDocument118 pagesStructure of MicrobesDeana Namirembe100% (6)
- Microbiology: PROKARYOTIC & EUKARYOTIC CELLSDocument10 pagesMicrobiology: PROKARYOTIC & EUKARYOTIC CELLSEmily RaeNo ratings yet
- Bacterial Disease AnalysisDocument63 pagesBacterial Disease AnalysisAshutoshkumar jhaNo ratings yet
- SHS - Biology 1 - Q1 - Module 3 - Reviewed - EditedDocument30 pagesSHS - Biology 1 - Q1 - Module 3 - Reviewed - EditedLiberty Salazar100% (2)
- BIO 275 - Chapter 3 and 4 QuestionsDocument3 pagesBIO 275 - Chapter 3 and 4 QuestionsLindsey StilleyNo ratings yet
- F. Y. B. Sc. (Microbiology) Paper-I Question Bank PDFDocument16 pagesF. Y. B. Sc. (Microbiology) Paper-I Question Bank PDFShipra ChakrabortyNo ratings yet
- Altas de Dasicladaceas P 1-50Document50 pagesAltas de Dasicladaceas P 1-508811791No ratings yet
- Comparing Extracellular Secretion Mechanisms in E. Coli Strains BL21 (DE3) and Nissle 1917Document10 pagesComparing Extracellular Secretion Mechanisms in E. Coli Strains BL21 (DE3) and Nissle 1917vikramreddy041302No ratings yet
- Soal ToeflDocument11 pagesSoal ToeflNurul A'la RojasNo ratings yet
- Chapter 4: ProtistaDocument8 pagesChapter 4: ProtistaNawal Che IsmailNo ratings yet
- Bio 1 PDFDocument13 pagesBio 1 PDFjayan sivaNo ratings yet
- What Are Cell OrganellesDocument19 pagesWhat Are Cell OrganellesMaraon CharitaNo ratings yet
- Important Questions For CBSE Class 11 Biology Chapter 8Document15 pagesImportant Questions For CBSE Class 11 Biology Chapter 8lavanya rajaNo ratings yet
- Anatomy of Bacteria: Morphology Structure FunctionDocument49 pagesAnatomy of Bacteria: Morphology Structure FunctionBlanche AltheaNo ratings yet
- Cell StructersDocument13 pagesCell StructersSumaiya JabinNo ratings yet
- Topic: Bacteria: EtymologyDocument28 pagesTopic: Bacteria: EtymologyVINDHYA SHANKERNo ratings yet
- Prokaryotic Profiles: The Bacteria and The Archaea: Week 2Document44 pagesProkaryotic Profiles: The Bacteria and The Archaea: Week 2Irwan M. IskoberNo ratings yet