Download as doc, pdf, or txt
You might also like
- Quiz 3 AnswersDocument3 pagesQuiz 3 AnswersEdilberto PerezNo ratings yet
- PBL SS 2 - Fetal DevelopmentDocument3 pagesPBL SS 2 - Fetal DevelopmentJellie MendozaNo ratings yet
- 3rd Lecture Morphological Changes During Maturation o Te GameteaDocument25 pages3rd Lecture Morphological Changes During Maturation o Te GameteaHussein Al Saedi100% (1)
- Animal DiversityDocument45 pagesAnimal DiversitySafiqa Tasfia100% (1)
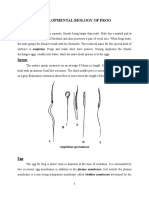
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
- Chapter 38 Animal DevelopmentDocument77 pagesChapter 38 Animal Developmentmaria banunaekNo ratings yet
- 8.introduction To EmbryologyDocument63 pages8.introduction To EmbryologyAhmed OrabyNo ratings yet
- Development of Frog Umanga Chapagain Read OnlyDocument9 pagesDevelopment of Frog Umanga Chapagain Read OnlySubarna PudasainiNo ratings yet
- Early DevelopmentDocument5 pagesEarly DevelopmentMarina of The SeaNo ratings yet
- GastrulationDocument1 pageGastrulationClaire BolalinNo ratings yet
- Early Development in BirdsDocument53 pagesEarly Development in BirdsMuqaddas NadeemNo ratings yet
- Cleavage: The Main Process of Animal DevelopmentDocument123 pagesCleavage: The Main Process of Animal DevelopmentNUR ASMA RIZKYNo ratings yet
- Lecture 16 Fate Mapping and Gastrulation in Sea UrchinDocument23 pagesLecture 16 Fate Mapping and Gastrulation in Sea UrchinIQra KanwalNo ratings yet
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Gastrulation in AmphibiansDocument20 pagesGastrulation in AmphibiansAnonymous mpD5IsJvaNo ratings yet
- Dichotomous KeynotesDocument19 pagesDichotomous KeynotesAnonymous vqO6zPNo ratings yet
- Metamorphism in InsectsDocument24 pagesMetamorphism in Insectsyayeg raja100% (1)
- 01 - Embryology (Theory)Document54 pages01 - Embryology (Theory)Simran Kashyap100% (1)
- Neurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes TheDocument6 pagesNeurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes Theaaliya saaheenNo ratings yet
- Rat Urinary and Reproductive SystemDocument4 pagesRat Urinary and Reproductive SystemAnkit NariyaNo ratings yet
- Introduction To Embryology 1Document72 pagesIntroduction To Embryology 1Sam MumoNo ratings yet
- ZL 124 Lecture 7 - Gastrulation in Birds & MammalsDocument36 pagesZL 124 Lecture 7 - Gastrulation in Birds & MammalsNestory MartineNo ratings yet
- Ex234.1 Ascaris Eggz PDFDocument7 pagesEx234.1 Ascaris Eggz PDFCarlo MendozaNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- DEVBIOL LE 1 ReviewerDocument43 pagesDEVBIOL LE 1 Reviewernamkimseo100% (1)
- Chapter 2 - Membrane PhysiologyDocument11 pagesChapter 2 - Membrane Physiologytahira UroojNo ratings yet
- Gastrulation LectureDocument74 pagesGastrulation LectureCynthiaNo ratings yet
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- IntroductionDocument24 pagesIntroductioninoka911No ratings yet
- Gametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalDocument16 pagesGametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalLim Yi QiNo ratings yet
- Animal Kingdom: Basis of ClassificationDocument15 pagesAnimal Kingdom: Basis of ClassificationChaitali Deshmukh100% (1)
- Population Growth CurveDocument16 pagesPopulation Growth CurvePRABIRA KUMAR SAHOONo ratings yet
- Chick GastrulationDocument6 pagesChick GastrulationAlyssa Gail de VeraNo ratings yet
- 4 - Cleavage and BlastulationDocument31 pages4 - Cleavage and BlastulationIRENE SEBASTIANNo ratings yet
- General Classification (All Phylum)Document16 pagesGeneral Classification (All Phylum)kiran PoudelNo ratings yet
- Male & Female Reproductive System: Dr. Zainab Neamat Al-TaeeDocument46 pagesMale & Female Reproductive System: Dr. Zainab Neamat Al-TaeeIn UsNo ratings yet
- 17 General Embryology MlabDocument44 pages17 General Embryology MlabJhu N EYNo ratings yet
- Muscular System Quiz Study GuideDocument10 pagesMuscular System Quiz Study Guidejbradee100% (2)
- Phylum Chordata: C O P C NDocument8 pagesPhylum Chordata: C O P C NBaikuntha SabarNo ratings yet
- Locomotion by Cilia and FlagellaDocument30 pagesLocomotion by Cilia and FlagellaSuchitra SharmaNo ratings yet
- EarthwormDocument23 pagesEarthwormNihal Jain100% (4)
- Excretion and HomeostasisDocument11 pagesExcretion and Homeostasisstarcandypricess100% (1)
- CH 26 Phylum PoriferaDocument51 pagesCH 26 Phylum Poriferaapi-244168124No ratings yet
- 4 Animal KingdomDocument33 pages4 Animal KingdomS SNo ratings yet
- Practical # 1serial Sections of FrogDocument36 pagesPractical # 1serial Sections of FrogMomina ButtNo ratings yet
- TOrsionDocument19 pagesTOrsionDrSneha VermaNo ratings yet
- Porifera MCQDocument4 pagesPorifera MCQRiwaj BogatiNo ratings yet
- Descriptive Embryology PDFDocument12 pagesDescriptive Embryology PDFMuhammad KhalilNo ratings yet
- Cytoplasmic InheritanceDocument13 pagesCytoplasmic InheritanceIzza NafisaNo ratings yet
- Complete Chordata NotesDocument99 pagesComplete Chordata NotesNarasimha MurthyNo ratings yet
- Lesson 11. Phylum Echinodermata PDFDocument37 pagesLesson 11. Phylum Echinodermata PDFJonard PedrosaNo ratings yet
- Notes On The Phylum PORIFERADocument20 pagesNotes On The Phylum PORIFERAnoblefx100% (1)
- Cell Parts & Functions TableDocument2 pagesCell Parts & Functions TableJerene BrownNo ratings yet
- Cell Growth and DivisionDocument15 pagesCell Growth and DivisionJohn Lester BerdinNo ratings yet
- Lecture 3 GametogenesisDocument28 pagesLecture 3 GametogenesisAparna PSNo ratings yet
- Lab Report Cell Bio MitosisDocument4 pagesLab Report Cell Bio Mitosisain syukriahNo ratings yet
- Rabbit RespirationDocument5 pagesRabbit RespirationshajehanNo ratings yet
- Sex Determination in DrosophilaDocument19 pagesSex Determination in DrosophilaSheerin Sulthana100% (2)
- Practical - DEVELOPMENTAL STAGES OF FROGDocument6 pagesPractical - DEVELOPMENTAL STAGES OF FROGtooba tahirNo ratings yet
- Frog EmbryologyDocument8 pagesFrog EmbryologyklumabanNo ratings yet
- Presentation Title: Your Company InformationDocument2 pagesPresentation Title: Your Company Informationangel_soshiNo ratings yet
- Presentation Title: Your Company InformationDocument2 pagesPresentation Title: Your Company Informationangel_soshiNo ratings yet
- R.T CreedDocument1 pageR.T Creedangel_soshiNo ratings yet
- How To Conduct A Semi Structured InterviewDocument3 pagesHow To Conduct A Semi Structured Interviewangel_soshiNo ratings yet
- Lesson 2 Parts of SpeechDocument18 pagesLesson 2 Parts of Speechangel_soshi100% (1)
- Rizal Family TreeDocument2 pagesRizal Family Treeangel_soshiNo ratings yet