Download as pdf or txt
You might also like
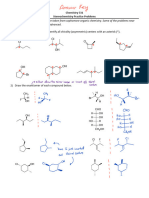
- 531 - Stereochem Practice KeyDocument4 pages531 - Stereochem Practice KeyAli MuhtashamNo ratings yet
- Lab Manual REEDocument60 pagesLab Manual REESHIVAM JHANo ratings yet
- Chemistry Lab Report 1212Document9 pagesChemistry Lab Report 1212api-284826801No ratings yet
- BUCX/il1EA!r Cabopi Fron Dinaluc: Lu Ka Ro EvarDocument16 pagesBUCX/il1EA!r Cabopi Fron Dinaluc: Lu Ka Ro EvartegavNo ratings yet
- Coat Color, Solar Heat Gain, And: Gre - MQ E. WalsbergDocument8 pagesCoat Color, Solar Heat Gain, And: Gre - MQ E. WalsbergRodrigo CarralNo ratings yet
- The Ultimate Infrared Handbook For R&D ProfessionalsDocument44 pagesThe Ultimate Infrared Handbook For R&D ProfessionalsIeremeiov Vladimir100% (5)
- Solar Radiation Resource Assessment by Means of Silicon CellsDocument9 pagesSolar Radiation Resource Assessment by Means of Silicon CellsalexanderritschelNo ratings yet
- Blip 1Document10 pagesBlip 1Dilusha SilvaNo ratings yet
- 8 Infra RedDocument41 pages8 Infra RedMykyl Dominic Duremdes LariosaNo ratings yet
- Wang Tao 1949Document26 pagesWang Tao 1949Kim AmnaNo ratings yet
- Applications of Thermal Imaging in Avian ScienceDocument13 pagesApplications of Thermal Imaging in Avian ScienceTinuszkaNo ratings yet
- One Picosecond Optical Nor Gate at Room Temperature With A Gaas Algaas Multiple Quantum Well Nonlinear Fabry-Perot ÉtalonDocument4 pagesOne Picosecond Optical Nor Gate at Room Temperature With A Gaas Algaas Multiple Quantum Well Nonlinear Fabry-Perot ÉtalonKadu BritoNo ratings yet
- The Geiger-Mueller Tube, Nuclear Electronics, and Counting StatisticsDocument4 pagesThe Geiger-Mueller Tube, Nuclear Electronics, and Counting StatisticsWalimatul FitriyahNo ratings yet
- Infrared Thermography On Ocular SurfaceDocument21 pagesInfrared Thermography On Ocular SurfaceEdson silvaNo ratings yet
- 1999 ArticleDocument8 pages1999 Articleelder diazNo ratings yet
- The American Mineralogist, VOL. 49, JULY - AUGUST' 1964Document16 pagesThe American Mineralogist, VOL. 49, JULY - AUGUST' 1964Quazai DailyNo ratings yet
- Solar Radiation and PyranometerDocument32 pagesSolar Radiation and PyranometerIrfan Khan100% (2)
- Thermal RadiationDocument5 pagesThermal RadiationBor OtNo ratings yet
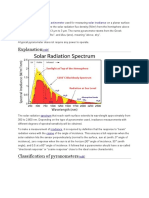
- Explanation: ῦ (pyr), meaning "fire", and νω ἄDocument6 pagesExplanation: ῦ (pyr), meaning "fire", and νω ἄMonika KshetriNo ratings yet
- Bolo MeterDocument6 pagesBolo MeterDemian PereiraNo ratings yet
- 1 s2.0 S0011227508001124 MainDocument6 pages1 s2.0 S0011227508001124 MainRev TamasNo ratings yet
- Solar Control Shading Devices 1958Document1 pageSolar Control Shading Devices 1958Batman WayneNo ratings yet
- 14 ApendicesDocument37 pages14 ApendicessorsanbarNo ratings yet
- Chromatic Cues To Trap The Oriental Fruit Fly, Bactrocera DorsalisDocument8 pagesChromatic Cues To Trap The Oriental Fruit Fly, Bactrocera Dorsalisgarcia31_2No ratings yet
- CollectorDocument5 pagesCollectorBranislavPetrovicNo ratings yet
- Pocket Guide Thermography enDocument56 pagesPocket Guide Thermography enMárton SolymosiNo ratings yet
- Camacho 1999Document9 pagesCamacho 1999Linggar T. GupitaNo ratings yet
- Atot-2008jtecha1100 1Document9 pagesAtot-2008jtecha1100 1Khalid AliNo ratings yet
- GLOSSARY of Thermography TermsDocument14 pagesGLOSSARY of Thermography TermsMohamed Al-OdatNo ratings yet
- Biophoton Emission: New Evidence For Coherence and DNA As SourceDocument20 pagesBiophoton Emission: New Evidence For Coherence and DNA As SourceValentina Carbini100% (1)
- Fairchild1966 PDFDocument17 pagesFairchild1966 PDFDiego Antonio Rodriguez CoronadoNo ratings yet
- Infrared Optical PyrometerDocument6 pagesInfrared Optical PyrometerinstrutechNo ratings yet
- TMPFCDB TMPDocument13 pagesTMPFCDB TMPFrontiersNo ratings yet
- G-Vs G-Vs G Vs G-Vs G VS: Building and Environment 177 (2020) 106875 F. Convertino Et AlDocument1 pageG-Vs G-Vs G Vs G-Vs G VS: Building and Environment 177 (2020) 106875 F. Convertino Et AlCovjece Ne Ljuti SeNo ratings yet
- EXPERIMENT 8. Monolayer Characterization: Contact Angles, Reflection Infrared Spectroscopy, and EllipsometryDocument9 pagesEXPERIMENT 8. Monolayer Characterization: Contact Angles, Reflection Infrared Spectroscopy, and EllipsometryavniNo ratings yet
- Infrared Thermography Principles and ApplicationsDocument10 pagesInfrared Thermography Principles and ApplicationsAlvaro ClarosNo ratings yet
- Anger Scitillation CameraDocument8 pagesAnger Scitillation CameraTuấn Hồ TrọngNo ratings yet
- Jtech1892 1Document13 pagesJtech1892 1R. SanchezNo ratings yet
- K. W. Geiger, Et Al., Slowing Down Spectrum and Neutron Temperature in A Thermal Neutron Neutron Flux Density StandardDocument6 pagesK. W. Geiger, Et Al., Slowing Down Spectrum and Neutron Temperature in A Thermal Neutron Neutron Flux Density Standard최재원No ratings yet
- CO2 Exchange of PlantsDocument10 pagesCO2 Exchange of PlantsAqsa AzeemNo ratings yet
- The Measurement of Absorptivity and Reflectivity: Ministry BF AviationDocument24 pagesThe Measurement of Absorptivity and Reflectivity: Ministry BF AviationCamilo Enrique Monroy CardenasNo ratings yet
- G. Sonnabend Et Al - High-Resolution Infrared Measurements of H2O and SiO in SunspotsDocument9 pagesG. Sonnabend Et Al - High-Resolution Infrared Measurements of H2O and SiO in SunspotsRemazzzNo ratings yet
- VHT PaintDocument11 pagesVHT PaintvytoNo ratings yet
- Response of The Lazio-Sirad Detector To Low Energy ElectronsDocument4 pagesResponse of The Lazio-Sirad Detector To Low Energy ElectronsMalikaNo ratings yet
- Tdepartment of Physiology, London,: Hospital CollegeDocument13 pagesTdepartment of Physiology, London,: Hospital Collegedesi trihartatiNo ratings yet
- Astm E408 1971Document5 pagesAstm E408 1971Hsaam HsaamNo ratings yet
- Desert Snails: Problems of Heat, Water and FoodDocument14 pagesDesert Snails: Problems of Heat, Water and FoodcoordinatedmultibeamsatelliteNo ratings yet
- Biophoton Emission: New Evidence For Coherence and DNA As SourceDocument20 pagesBiophoton Emission: New Evidence For Coherence and DNA As SourceLurzizareNo ratings yet
- Temperature MeasurementDocument9 pagesTemperature Measurementbarry nancooNo ratings yet
- Mer016 922..930Document9 pagesMer016 922..930kshazadNo ratings yet
- TL and OSLDocument18 pagesTL and OSLRaj PrakashNo ratings yet
- ACETANILIDEDocument10 pagesACETANILIDEDiego AlonsoNo ratings yet
- Infrared Pyrometer HandbookDocument19 pagesInfrared Pyrometer Handbookjoyous leeNo ratings yet
- Satellite Remote Sensing of The Environment: Introduction and BackgroundDocument10 pagesSatellite Remote Sensing of The Environment: Introduction and BackgrounddejenabNo ratings yet
- 09 Chapter 2Document15 pages09 Chapter 2kohonNo ratings yet
- SolarDocument13 pagesSolarVinay Maisuriya0% (1)
- Radiation of Heat From Human BodyDocument15 pagesRadiation of Heat From Human BodyrocguedesNo ratings yet
- Self Calibrating Infrared Thermometer FoDocument6 pagesSelf Calibrating Infrared Thermometer Fomarcelo.lacknerNo ratings yet
- Journal 10Document14 pagesJournal 10ZulAmNo ratings yet
- Effects of UV Radiation On Photosynthesis of Phytoplankton Exposed To Solar Simulator LightDocument8 pagesEffects of UV Radiation On Photosynthesis of Phytoplankton Exposed To Solar Simulator LightAlexsandro ClaudinoNo ratings yet
- Radiation 5 PDF FreeDocument18 pagesRadiation 5 PDF FreeRose RayNo ratings yet
- Tariro Moto Project Proposal UpdatedDocument13 pagesTariro Moto Project Proposal Updatedtapiwa guy nyamukapaNo ratings yet
- 2 3 Atomic ModelsDocument19 pages2 3 Atomic Modelsapi-259040408No ratings yet
- TAAMPP Research Summary PDFDocument14 pagesTAAMPP Research Summary PDFBaltranslNo ratings yet
- G9 2017 2018Document9 pagesG9 2017 2018Fatima SlimNo ratings yet
- 02-MSc Grain Preservation and ProcessingDocument761 pages02-MSc Grain Preservation and Processing11751175No ratings yet
- Comparison of EN ISO 15614-1 - 2012 Vs 2017Document6 pagesComparison of EN ISO 15614-1 - 2012 Vs 2017Swiss CIS AGNo ratings yet
- Astm d1785Document5 pagesAstm d1785Kehinde AdebayoNo ratings yet
- Structural Stability 1Document10 pagesStructural Stability 1Tugas Hutomo PutraNo ratings yet
- 1457156375-Inorganic Chemistry Revision Book JEE Main-2016.PmdDocument49 pages1457156375-Inorganic Chemistry Revision Book JEE Main-2016.PmdSummer GreenNo ratings yet
- RM 732 735 DesinfeccionDocument2 pagesRM 732 735 DesinfeccionCesar David RengifoNo ratings yet
- CP Offshore Anode CalculationDocument3 pagesCP Offshore Anode CalculationmaniechemistNo ratings yet
- Coil Oscillator Trigger Circuit Output Circuit Motion Direction Proximity Switch TargetDocument14 pagesCoil Oscillator Trigger Circuit Output Circuit Motion Direction Proximity Switch Targetrc_casas99No ratings yet
- Re64354 2016-04Document16 pagesRe64354 2016-04br1404No ratings yet
- HDPE Pipes Fittings PDFDocument56 pagesHDPE Pipes Fittings PDFInthees RagavanNo ratings yet
- Hilti - Anchor Fastening Technology ManualDocument0 pagesHilti - Anchor Fastening Technology ManualDavid Salomão Pinto Castanho BizarroNo ratings yet
- Moiusture Equilibrium and Moisture Migration Within Transforme Insulation SystemDocument53 pagesMoiusture Equilibrium and Moisture Migration Within Transforme Insulation SystemDiegoDonosoNo ratings yet
- Science Grade 7 Test BankDocument3 pagesScience Grade 7 Test BankAlisha EnguanchoNo ratings yet
- Sample Size and Meaningful Gold AnalyisDocument20 pagesSample Size and Meaningful Gold AnalyisDash WeeksNo ratings yet
- Mtech Thesis NirliptaDocument83 pagesMtech Thesis NirliptaBiswajit MohantyNo ratings yet
- GTL-81-01 Rev 03 Protective CoatingsDocument36 pagesGTL-81-01 Rev 03 Protective CoatingsPurush Purush100% (1)
- UCD 115 Instrumental Analysis: UC DavisDocument73 pagesUCD 115 Instrumental Analysis: UC Daviskhanh khanhNo ratings yet
- 07 4 2017123Document111 pages07 4 2017123RajeshYadav100% (1)
- GSP Test For AD Post, 2014Document5 pagesGSP Test For AD Post, 2014Tahir Khattak100% (2)
- NinhydrinDocument5 pagesNinhydrinMahboob Iqbal0% (1)
- Previous HSE Questions and Answers of The Chapter "SOLUTIONS"Document97 pagesPrevious HSE Questions and Answers of The Chapter "SOLUTIONS"ABDUL RAZACKNo ratings yet
- Diagramas TT de Ferro e Aço - Aço de Alta ResistênciaDocument111 pagesDiagramas TT de Ferro e Aço - Aço de Alta ResistênciaFernando Venceslau100% (1)
- Detailed Syllabus of Soil MechanicsDocument9 pagesDetailed Syllabus of Soil MechanicsRajesh KhadkaNo ratings yet
- An Insight Into PlasticsDocument7 pagesAn Insight Into PlasticsVivek AradhyaNo ratings yet