
Malaria 03
Malaria 03
You might also like
- Schizophrenia Case StudyDocument13 pagesSchizophrenia Case StudyAnonymous Hfrl594No ratings yet
- Hema RecallDocument11 pagesHema Recallmkct111100% (1)
- Excerpt: "Fat Chance" by Robert LustigDocument3 pagesExcerpt: "Fat Chance" by Robert Lustigwamu885067% (3)
- 020147Document9 pages020147Shital BishtNo ratings yet
- David Masopust Sang-Jun Ha Vaiva Vezys and Rafi Ahmed Stimulation History Dictates Memory CD8 T Cell PehnotypeDocument10 pagesDavid Masopust Sang-Jun Ha Vaiva Vezys and Rafi Ahmed Stimulation History Dictates Memory CD8 T Cell PehnotypeRepositorioNo ratings yet
- R 2017 - Regulatory T Cells in Cancer ImmunotherapyDocument10 pagesR 2017 - Regulatory T Cells in Cancer Immunotherapy1262615286No ratings yet
- Molecular ImmunologyDocument12 pagesMolecular ImmunologymmsNo ratings yet
- 157549.2 20220517120354 CoveredDocument15 pages157549.2 20220517120354 CoveredJulián DuránNo ratings yet
- Regulatory T Cells - How Do They Suppress Immine ResponsesDocument7 pagesRegulatory T Cells - How Do They Suppress Immine ResponsesWendi IochNo ratings yet
- Ilar 46 3 230Document11 pagesIlar 46 3 230Rosydina Robi'aqolbiNo ratings yet
- Modeling Human Adaptive Immune Responses With Tonsil OrganoidsDocument27 pagesModeling Human Adaptive Immune Responses With Tonsil Organoidsissis alppNo ratings yet
- Antigen-Presentation and The Role of Dendritic Cells in PeriodontitisDocument23 pagesAntigen-Presentation and The Role of Dendritic Cells in PeriodontitiskochikaghochiNo ratings yet
- KPC Mouse ModelDocument16 pagesKPC Mouse Modeljoe smoNo ratings yet
- Monophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andDocument14 pagesMonophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andLeandro CarreñoNo ratings yet
- Transialidasa ISPA 2017 PDFDocument18 pagesTransialidasa ISPA 2017 PDFVic PastorNo ratings yet
- Age and ImmunityDocument6 pagesAge and ImmunityGene Ryuzaki SeseNo ratings yet
- Reviews: T Cell-Based Therapies: Challenges and PerspectivesDocument15 pagesReviews: T Cell-Based Therapies: Challenges and PerspectivesDaniel Luis Meléndez ChoqueNo ratings yet
- T CellsDocument12 pagesT CellsGabriela CismaruNo ratings yet
- The Role of Immune Checkpoints in Immunity and CancerDocument8 pagesThe Role of Immune Checkpoints in Immunity and Cancernikos tsatsakisNo ratings yet
- Mouse ModelsDocument32 pagesMouse ModelsCiocan AlexandraNo ratings yet
- J Immunol 2010 Mueller 2182 90Document10 pagesJ Immunol 2010 Mueller 2182 90winsarkarNo ratings yet
- PHD Thesis Dendritic CellsDocument5 pagesPHD Thesis Dendritic Cellsashleyjonesmobile100% (1)
- Royal JellyDocument10 pagesRoyal JellyninukNo ratings yet
- 1 s2.0 S1074761316304009 MainDocument18 pages1 s2.0 S1074761316304009 MainfranciscaNo ratings yet
- AJPS Volume 64 Issue 2 Pages 41-68Document28 pagesAJPS Volume 64 Issue 2 Pages 41-68Mohamad MostafaNo ratings yet
- The Bone Marrow Protects and OptimizesImmunological Memory During Dietary RestrictionDocument30 pagesThe Bone Marrow Protects and OptimizesImmunological Memory During Dietary RestrictionAbraao 2000No ratings yet
- Nov 151Document6 pagesNov 151alexandre06frNo ratings yet
- Tamil Nadu Class 12 - Bio-Zoology-Zoology - English Medium - Possible 5 Mark Questions With Answer.Document34 pagesTamil Nadu Class 12 - Bio-Zoology-Zoology - English Medium - Possible 5 Mark Questions With Answer.revamanian57% (7)
- The FEBS Journal - 2021 - Zhang - Hallmarks of The Aging T Cell SystemDocument20 pagesThe FEBS Journal - 2021 - Zhang - Hallmarks of The Aging T Cell SystemSubhajit DuttaNo ratings yet
- 1 s2.0 S0145305X1100303X MainDocument13 pages1 s2.0 S0145305X1100303X MainEdu AlmeidaNo ratings yet
- Secreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNDocument10 pagesSecreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNEdy NoveryNo ratings yet
- Seminario 11 InmunoDocument16 pagesSeminario 11 InmunoBrenda Baltodano GarcìaNo ratings yet
- Cluster of Differentiation Antigens: Essential Roles in The Identification of Teleost Fish T LymphocytesDocument14 pagesCluster of Differentiation Antigens: Essential Roles in The Identification of Teleost Fish T LymphocytesBayuNo ratings yet
- Immune Response and Immunotherapy in Chronic Lymphocytic LeukemiaDocument25 pagesImmune Response and Immunotherapy in Chronic Lymphocytic Leukemiakj185No ratings yet
- Cells: Regulatory T Cell-Enhancing Therapies To Treat AtherosclerosisDocument17 pagesCells: Regulatory T Cell-Enhancing Therapies To Treat AtherosclerosisLAURA VALENTINA CASTAÑEDA TRUJILLONo ratings yet
- Envhper00522 0014Document5 pagesEnvhper00522 0014Afaq AhmadNo ratings yet
- Defense Systems in FishDocument38 pagesDefense Systems in FishEddyNo ratings yet
- Articulo Innata 2002Document21 pagesArticulo Innata 2002Diego Omar Perez CardenasNo ratings yet
- Immuno Senescence PublishedDocument5 pagesImmuno Senescence PublishedAcengHamudinNo ratings yet
- Text Full LifeDocument9 pagesText Full Lifezurique32No ratings yet
- Dendritic Cell Gene TherapyDocument16 pagesDendritic Cell Gene TherapyazharbattooNo ratings yet
- Vitd TregDocument8 pagesVitd Tregervina 119260026No ratings yet
- tmp536C TMPDocument11 pagestmp536C TMPFrontiersNo ratings yet
- Avances en Nuevas Estrategias de Vacunas para La Inmunoterapia y Prevención Del Cáncer JAY A. BERZOFSKY 2004Document11 pagesAvances en Nuevas Estrategias de Vacunas para La Inmunoterapia y Prevención Del Cáncer JAY A. BERZOFSKY 2004Ramiro J. Rodriguez GarciaNo ratings yet
- Alternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalDocument7 pagesAlternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalTruc PhamNo ratings yet
- In Silico Leishmania Major in VitroDocument13 pagesIn Silico Leishmania Major in VitroshahabNo ratings yet
- Cellular Innate Immunity: An Old Game With New PlayersDocument15 pagesCellular Innate Immunity: An Old Game With New Playersklara kapitan08No ratings yet
- Tumor XenograftDocument3 pagesTumor Xenograft2jbkrf5bkcNo ratings yet
- 3 Pal C Et Al 2007 Indian J Exp BiolDocument14 pages3 Pal C Et Al 2007 Indian J Exp BiolDipyaman GangulyNo ratings yet
- Binding of The Fap2 Protein of Fusobacterium Nucleatum To Human Inhibitory Receptor TIGIT Protects Tumors From Immune Cell Attack 2015 ImmunityDocument13 pagesBinding of The Fap2 Protein of Fusobacterium Nucleatum To Human Inhibitory Receptor TIGIT Protects Tumors From Immune Cell Attack 2015 ImmunityMatthew MacowanNo ratings yet
- Immunomodulatory Properties of Dental Tissue-Derived Mesenchymal Stem CellsDocument25 pagesImmunomodulatory Properties of Dental Tissue-Derived Mesenchymal Stem CellsFernando CruzNo ratings yet
- Kobezda Et Al-T Cells in Mice and Men-NRR-05.08.2013Document29 pagesKobezda Et Al-T Cells in Mice and Men-NRR-05.08.2013Anett Pappné LeppNo ratings yet
- A Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesDocument19 pagesA Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesIJAR JOURNALNo ratings yet
- Cytokines: Names and Numbers You Should Care About: Stephen R. Holdsworth and Poh-Yi GanDocument12 pagesCytokines: Names and Numbers You Should Care About: Stephen R. Holdsworth and Poh-Yi GanKrisztián TóthNo ratings yet
- Part 2 Basics of ImmunologyDocument6 pagesPart 2 Basics of Immunologyabc abcNo ratings yet
- Dendritic Cell-Based Cancer VaccinesDocument8 pagesDendritic Cell-Based Cancer VaccinesJamil ANo ratings yet
- CTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceDocument10 pagesCTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceMaxime PorcoNo ratings yet
- 3493 FullDocument8 pages3493 Fullgaa5No ratings yet
- TIM Genes - A Family of Cell Surface Phosphatidylserine Receptors That Regulate Innate and Adaptive ImmunityDocument18 pagesTIM Genes - A Family of Cell Surface Phosphatidylserine Receptors That Regulate Innate and Adaptive ImmunityJun LiNo ratings yet
- Sikasunge Et Al. 2009Document9 pagesSikasunge Et Al. 2009Luz Cielo Manrique CordovaNo ratings yet
- 8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlDocument12 pages8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlSage RadachowskyNo ratings yet
- IDO1Document13 pagesIDO1Victor HaidarNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Alkaloids IntroductionDocument35 pagesAlkaloids Introductionalexpharm0% (1)
- ForteBio App Note 1Document2 pagesForteBio App Note 1Fernando OviedoNo ratings yet
- CollagenDocument18 pagesCollagenAhsan AliNo ratings yet
- 1 s2.0 S2666765722001661 MainDocument11 pages1 s2.0 S2666765722001661 MainJoshua HammedNo ratings yet
- Massachusetts Department of Public Health COVID-19 DashboardDocument24 pagesMassachusetts Department of Public Health COVID-19 DashboardJohn WallerNo ratings yet
- NBME CompDocument7 pagesNBME CompWinter Ohio100% (4)
- Electrochemical Methods For The Analysis of Clinically RelevantDocument90 pagesElectrochemical Methods For The Analysis of Clinically Relevantch_ymyaaNo ratings yet
- Zoo Vert Molecular Techniques For Sex Identification of Captive BirdsDocument9 pagesZoo Vert Molecular Techniques For Sex Identification of Captive BirdsFelipe AraújoNo ratings yet
- The Brain Slice TechniqueDocument3 pagesThe Brain Slice Techniqueapi-3846255No ratings yet
- Pediatricobesity: Etiologyandtreatment: Melissa K. Crocker,, Jack A. YanovskiDocument24 pagesPediatricobesity: Etiologyandtreatment: Melissa K. Crocker,, Jack A. YanovskiAyline Araceli AlavaNo ratings yet
- Exp11 PDFDocument6 pagesExp11 PDFCloudCrescentNo ratings yet
- DNA Barcoding and Molecular PhylogenyDocument431 pagesDNA Barcoding and Molecular PhylogenyRoxana Diana GherghelucaNo ratings yet
- 11 Biology Notes ch19 Excretory Products and Their Elimination PDFDocument3 pages11 Biology Notes ch19 Excretory Products and Their Elimination PDFRamachandranPerumalNo ratings yet
- BiochemDocument2 pagesBiochemHimashree DasNo ratings yet
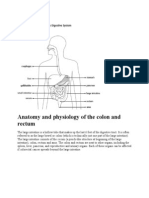
- Anatomy and Physiology of The Colon and RectumDocument4 pagesAnatomy and Physiology of The Colon and RectumGeraldine Gallaron - CasipongNo ratings yet
- Uts Reviewer Cover To CoverDocument19 pagesUts Reviewer Cover To Covermyungsoo100% (2)
- A Sonographic Approach To The Prenatal Diagnosis of Skeletal DysplasiasDocument19 pagesA Sonographic Approach To The Prenatal Diagnosis of Skeletal DysplasiasminhhaiNo ratings yet
- EEG LabDocument13 pagesEEG LabMingmo_Lee_9319No ratings yet
- sk-gupta-biochemistry-pdf-14Document2 pagessk-gupta-biochemistry-pdf-14kamleshmeena2126No ratings yet
- Classification of Veneer PreparationsDocument11 pagesClassification of Veneer PreparationsVinisha Vipin SharmaNo ratings yet
- Protein SynthesisDocument29 pagesProtein SynthesisEugenia Migranova100% (9)
- Teratogenesis and Its Role in DevlopmentDocument16 pagesTeratogenesis and Its Role in DevlopmentmmNo ratings yet
- Signs and Symptoms of AdhdDocument7 pagesSigns and Symptoms of AdhdKUNNAMPALLIL GEJO JOHNNo ratings yet
- Early HumanDocument4 pagesEarly HumanPeem JatabutNo ratings yet
- Pre-Dent Press 8.5Document8 pagesPre-Dent Press 8.5Corissa ChangNo ratings yet
- Organization and MGT of Sports EventsDocument31 pagesOrganization and MGT of Sports Eventsjer ricNo ratings yet
- Stop AIDS. Keep The Promise.: L'Automédicat-ion Au Liban NordDocument13 pagesStop AIDS. Keep The Promise.: L'Automédicat-ion Au Liban Nordhussein bitarNo ratings yet
























































































Download as pdf or txt
You might also like
- Schizophrenia Case StudyDocument13 pagesSchizophrenia Case StudyAnonymous Hfrl594No ratings yet
- Hema RecallDocument11 pagesHema Recallmkct111100% (1)
- Excerpt: "Fat Chance" by Robert LustigDocument3 pagesExcerpt: "Fat Chance" by Robert Lustigwamu885067% (3)
- 020147Document9 pages020147Shital BishtNo ratings yet
- David Masopust Sang-Jun Ha Vaiva Vezys and Rafi Ahmed Stimulation History Dictates Memory CD8 T Cell PehnotypeDocument10 pagesDavid Masopust Sang-Jun Ha Vaiva Vezys and Rafi Ahmed Stimulation History Dictates Memory CD8 T Cell PehnotypeRepositorioNo ratings yet
- R 2017 - Regulatory T Cells in Cancer ImmunotherapyDocument10 pagesR 2017 - Regulatory T Cells in Cancer Immunotherapy1262615286No ratings yet
- Molecular ImmunologyDocument12 pagesMolecular ImmunologymmsNo ratings yet
- 157549.2 20220517120354 CoveredDocument15 pages157549.2 20220517120354 CoveredJulián DuránNo ratings yet
- Regulatory T Cells - How Do They Suppress Immine ResponsesDocument7 pagesRegulatory T Cells - How Do They Suppress Immine ResponsesWendi IochNo ratings yet
- Ilar 46 3 230Document11 pagesIlar 46 3 230Rosydina Robi'aqolbiNo ratings yet
- Modeling Human Adaptive Immune Responses With Tonsil OrganoidsDocument27 pagesModeling Human Adaptive Immune Responses With Tonsil Organoidsissis alppNo ratings yet
- Antigen-Presentation and The Role of Dendritic Cells in PeriodontitisDocument23 pagesAntigen-Presentation and The Role of Dendritic Cells in PeriodontitiskochikaghochiNo ratings yet
- KPC Mouse ModelDocument16 pagesKPC Mouse Modeljoe smoNo ratings yet
- Monophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andDocument14 pagesMonophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andLeandro CarreñoNo ratings yet
- Transialidasa ISPA 2017 PDFDocument18 pagesTransialidasa ISPA 2017 PDFVic PastorNo ratings yet
- Age and ImmunityDocument6 pagesAge and ImmunityGene Ryuzaki SeseNo ratings yet
- Reviews: T Cell-Based Therapies: Challenges and PerspectivesDocument15 pagesReviews: T Cell-Based Therapies: Challenges and PerspectivesDaniel Luis Meléndez ChoqueNo ratings yet
- T CellsDocument12 pagesT CellsGabriela CismaruNo ratings yet
- The Role of Immune Checkpoints in Immunity and CancerDocument8 pagesThe Role of Immune Checkpoints in Immunity and Cancernikos tsatsakisNo ratings yet
- Mouse ModelsDocument32 pagesMouse ModelsCiocan AlexandraNo ratings yet
- J Immunol 2010 Mueller 2182 90Document10 pagesJ Immunol 2010 Mueller 2182 90winsarkarNo ratings yet
- PHD Thesis Dendritic CellsDocument5 pagesPHD Thesis Dendritic Cellsashleyjonesmobile100% (1)
- Royal JellyDocument10 pagesRoyal JellyninukNo ratings yet
- 1 s2.0 S1074761316304009 MainDocument18 pages1 s2.0 S1074761316304009 MainfranciscaNo ratings yet
- AJPS Volume 64 Issue 2 Pages 41-68Document28 pagesAJPS Volume 64 Issue 2 Pages 41-68Mohamad MostafaNo ratings yet
- The Bone Marrow Protects and OptimizesImmunological Memory During Dietary RestrictionDocument30 pagesThe Bone Marrow Protects and OptimizesImmunological Memory During Dietary RestrictionAbraao 2000No ratings yet
- Nov 151Document6 pagesNov 151alexandre06frNo ratings yet
- Tamil Nadu Class 12 - Bio-Zoology-Zoology - English Medium - Possible 5 Mark Questions With Answer.Document34 pagesTamil Nadu Class 12 - Bio-Zoology-Zoology - English Medium - Possible 5 Mark Questions With Answer.revamanian57% (7)
- The FEBS Journal - 2021 - Zhang - Hallmarks of The Aging T Cell SystemDocument20 pagesThe FEBS Journal - 2021 - Zhang - Hallmarks of The Aging T Cell SystemSubhajit DuttaNo ratings yet
- 1 s2.0 S0145305X1100303X MainDocument13 pages1 s2.0 S0145305X1100303X MainEdu AlmeidaNo ratings yet
- Secreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNDocument10 pagesSecreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNEdy NoveryNo ratings yet
- Seminario 11 InmunoDocument16 pagesSeminario 11 InmunoBrenda Baltodano GarcìaNo ratings yet
- Cluster of Differentiation Antigens: Essential Roles in The Identification of Teleost Fish T LymphocytesDocument14 pagesCluster of Differentiation Antigens: Essential Roles in The Identification of Teleost Fish T LymphocytesBayuNo ratings yet
- Immune Response and Immunotherapy in Chronic Lymphocytic LeukemiaDocument25 pagesImmune Response and Immunotherapy in Chronic Lymphocytic Leukemiakj185No ratings yet
- Cells: Regulatory T Cell-Enhancing Therapies To Treat AtherosclerosisDocument17 pagesCells: Regulatory T Cell-Enhancing Therapies To Treat AtherosclerosisLAURA VALENTINA CASTAÑEDA TRUJILLONo ratings yet
- Envhper00522 0014Document5 pagesEnvhper00522 0014Afaq AhmadNo ratings yet
- Defense Systems in FishDocument38 pagesDefense Systems in FishEddyNo ratings yet
- Articulo Innata 2002Document21 pagesArticulo Innata 2002Diego Omar Perez CardenasNo ratings yet
- Immuno Senescence PublishedDocument5 pagesImmuno Senescence PublishedAcengHamudinNo ratings yet
- Text Full LifeDocument9 pagesText Full Lifezurique32No ratings yet
- Dendritic Cell Gene TherapyDocument16 pagesDendritic Cell Gene TherapyazharbattooNo ratings yet
- Vitd TregDocument8 pagesVitd Tregervina 119260026No ratings yet
- tmp536C TMPDocument11 pagestmp536C TMPFrontiersNo ratings yet
- Avances en Nuevas Estrategias de Vacunas para La Inmunoterapia y Prevención Del Cáncer JAY A. BERZOFSKY 2004Document11 pagesAvances en Nuevas Estrategias de Vacunas para La Inmunoterapia y Prevención Del Cáncer JAY A. BERZOFSKY 2004Ramiro J. Rodriguez GarciaNo ratings yet
- Alternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalDocument7 pagesAlternative Immunomodulatory Strategies For Xenotransplantation: CD80/CD86-CTLA4 Pathway-Modified Immature Dendritic Cells Promote Xenograft SurvivalTruc PhamNo ratings yet
- In Silico Leishmania Major in VitroDocument13 pagesIn Silico Leishmania Major in VitroshahabNo ratings yet
- Cellular Innate Immunity: An Old Game With New PlayersDocument15 pagesCellular Innate Immunity: An Old Game With New Playersklara kapitan08No ratings yet
- Tumor XenograftDocument3 pagesTumor Xenograft2jbkrf5bkcNo ratings yet
- 3 Pal C Et Al 2007 Indian J Exp BiolDocument14 pages3 Pal C Et Al 2007 Indian J Exp BiolDipyaman GangulyNo ratings yet
- Binding of The Fap2 Protein of Fusobacterium Nucleatum To Human Inhibitory Receptor TIGIT Protects Tumors From Immune Cell Attack 2015 ImmunityDocument13 pagesBinding of The Fap2 Protein of Fusobacterium Nucleatum To Human Inhibitory Receptor TIGIT Protects Tumors From Immune Cell Attack 2015 ImmunityMatthew MacowanNo ratings yet
- Immunomodulatory Properties of Dental Tissue-Derived Mesenchymal Stem CellsDocument25 pagesImmunomodulatory Properties of Dental Tissue-Derived Mesenchymal Stem CellsFernando CruzNo ratings yet
- Kobezda Et Al-T Cells in Mice and Men-NRR-05.08.2013Document29 pagesKobezda Et Al-T Cells in Mice and Men-NRR-05.08.2013Anett Pappné LeppNo ratings yet
- A Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesDocument19 pagesA Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesIJAR JOURNALNo ratings yet
- Cytokines: Names and Numbers You Should Care About: Stephen R. Holdsworth and Poh-Yi GanDocument12 pagesCytokines: Names and Numbers You Should Care About: Stephen R. Holdsworth and Poh-Yi GanKrisztián TóthNo ratings yet
- Part 2 Basics of ImmunologyDocument6 pagesPart 2 Basics of Immunologyabc abcNo ratings yet
- Dendritic Cell-Based Cancer VaccinesDocument8 pagesDendritic Cell-Based Cancer VaccinesJamil ANo ratings yet
- CTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceDocument10 pagesCTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceMaxime PorcoNo ratings yet
- 3493 FullDocument8 pages3493 Fullgaa5No ratings yet
- TIM Genes - A Family of Cell Surface Phosphatidylserine Receptors That Regulate Innate and Adaptive ImmunityDocument18 pagesTIM Genes - A Family of Cell Surface Phosphatidylserine Receptors That Regulate Innate and Adaptive ImmunityJun LiNo ratings yet
- Sikasunge Et Al. 2009Document9 pagesSikasunge Et Al. 2009Luz Cielo Manrique CordovaNo ratings yet
- 8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlDocument12 pages8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlSage RadachowskyNo ratings yet
- IDO1Document13 pagesIDO1Victor HaidarNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Alkaloids IntroductionDocument35 pagesAlkaloids Introductionalexpharm0% (1)
- ForteBio App Note 1Document2 pagesForteBio App Note 1Fernando OviedoNo ratings yet
- CollagenDocument18 pagesCollagenAhsan AliNo ratings yet
- 1 s2.0 S2666765722001661 MainDocument11 pages1 s2.0 S2666765722001661 MainJoshua HammedNo ratings yet
- Massachusetts Department of Public Health COVID-19 DashboardDocument24 pagesMassachusetts Department of Public Health COVID-19 DashboardJohn WallerNo ratings yet
- NBME CompDocument7 pagesNBME CompWinter Ohio100% (4)
- Electrochemical Methods For The Analysis of Clinically RelevantDocument90 pagesElectrochemical Methods For The Analysis of Clinically Relevantch_ymyaaNo ratings yet
- Zoo Vert Molecular Techniques For Sex Identification of Captive BirdsDocument9 pagesZoo Vert Molecular Techniques For Sex Identification of Captive BirdsFelipe AraújoNo ratings yet
- The Brain Slice TechniqueDocument3 pagesThe Brain Slice Techniqueapi-3846255No ratings yet
- Pediatricobesity: Etiologyandtreatment: Melissa K. Crocker,, Jack A. YanovskiDocument24 pagesPediatricobesity: Etiologyandtreatment: Melissa K. Crocker,, Jack A. YanovskiAyline Araceli AlavaNo ratings yet
- Exp11 PDFDocument6 pagesExp11 PDFCloudCrescentNo ratings yet
- DNA Barcoding and Molecular PhylogenyDocument431 pagesDNA Barcoding and Molecular PhylogenyRoxana Diana GherghelucaNo ratings yet
- 11 Biology Notes ch19 Excretory Products and Their Elimination PDFDocument3 pages11 Biology Notes ch19 Excretory Products and Their Elimination PDFRamachandranPerumalNo ratings yet
- BiochemDocument2 pagesBiochemHimashree DasNo ratings yet
- Anatomy and Physiology of The Colon and RectumDocument4 pagesAnatomy and Physiology of The Colon and RectumGeraldine Gallaron - CasipongNo ratings yet
- Uts Reviewer Cover To CoverDocument19 pagesUts Reviewer Cover To Covermyungsoo100% (2)
- A Sonographic Approach To The Prenatal Diagnosis of Skeletal DysplasiasDocument19 pagesA Sonographic Approach To The Prenatal Diagnosis of Skeletal DysplasiasminhhaiNo ratings yet
- EEG LabDocument13 pagesEEG LabMingmo_Lee_9319No ratings yet
- sk-gupta-biochemistry-pdf-14Document2 pagessk-gupta-biochemistry-pdf-14kamleshmeena2126No ratings yet
- Classification of Veneer PreparationsDocument11 pagesClassification of Veneer PreparationsVinisha Vipin SharmaNo ratings yet
- Protein SynthesisDocument29 pagesProtein SynthesisEugenia Migranova100% (9)
- Teratogenesis and Its Role in DevlopmentDocument16 pagesTeratogenesis and Its Role in DevlopmentmmNo ratings yet
- Signs and Symptoms of AdhdDocument7 pagesSigns and Symptoms of AdhdKUNNAMPALLIL GEJO JOHNNo ratings yet
- Early HumanDocument4 pagesEarly HumanPeem JatabutNo ratings yet
- Pre-Dent Press 8.5Document8 pagesPre-Dent Press 8.5Corissa ChangNo ratings yet
- Organization and MGT of Sports EventsDocument31 pagesOrganization and MGT of Sports Eventsjer ricNo ratings yet
- Stop AIDS. Keep The Promise.: L'Automédicat-ion Au Liban NordDocument13 pagesStop AIDS. Keep The Promise.: L'Automédicat-ion Au Liban Nordhussein bitarNo ratings yet