Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5834)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (824)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (405)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Riseberga Park - Restoration Project-1Document44 pagesRiseberga Park - Restoration Project-1TomNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Garden Design by Chris YoungDocument362 pagesGarden Design by Chris Younghadi100% (6)
- Biogeography BookDocument72 pagesBiogeography BookAnkit Anand0% (1)
- Ranald Lawrence Building On The HorizonDocument28 pagesRanald Lawrence Building On The HorizonCsenge FülöpNo ratings yet
- Barile&Brandon (Eds) - Household Chores and Household Choices Theoretizing The Domestic Sphere in Historical Archaeology PDFDocument330 pagesBarile&Brandon (Eds) - Household Chores and Household Choices Theoretizing The Domestic Sphere in Historical Archaeology PDFNancyNo ratings yet
- Soil Improvement Technique and LimitationsDocument4 pagesSoil Improvement Technique and LimitationsMuhammad SyafiqNo ratings yet
- Hugelkultur: The Ultimate Raised Garden BedsDocument23 pagesHugelkultur: The Ultimate Raised Garden BedsDaniel Pop100% (2)
- Production Technology of TobaccoDocument30 pagesProduction Technology of TobaccoWaqas AhmadNo ratings yet
- Hortuseum. Musealisation of The European Gardens in The Twenty-First CenturyDocument68 pagesHortuseum. Musealisation of The European Gardens in The Twenty-First CenturyJacek KuśmierskiNo ratings yet
- Project 2Document5 pagesProject 2api-302630382No ratings yet
- Science 1 - Q4 - L1 - Earth and Its Natural Resources - LandformsDocument11 pagesScience 1 - Q4 - L1 - Earth and Its Natural Resources - LandformsAira Shane LladaNo ratings yet
- 3 SennaDocument11 pages3 SennajayNo ratings yet
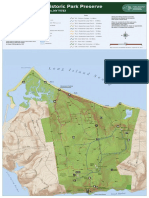
- Caumsett State Historic Park Preserve: Isla ND SoDocument1 pageCaumsett State Historic Park Preserve: Isla ND SoJosh WexlerNo ratings yet
- Lightweight Roof Garden GuidelinesDocument3 pagesLightweight Roof Garden GuidelinesSabiq ZinNo ratings yet
- Alluvial FanDocument5 pagesAlluvial FanAnonymous QEcQfTeHlNo ratings yet
- Runoff Estimation, and Surface Erosion and ControlDocument76 pagesRunoff Estimation, and Surface Erosion and ControlVivek KasarNo ratings yet
- Japanese Black Pine JBP - GraftingDocument3 pagesJapanese Black Pine JBP - GraftingManoj KumarNo ratings yet
- 205 459 1 SMDocument13 pages205 459 1 SMFakhriAnwarShenxoNo ratings yet
- Savanna: Navigation Search Savannah (Disambiguation)Document7 pagesSavanna: Navigation Search Savannah (Disambiguation)sNo ratings yet
- Politeknik Negeri Medan: Laboratorium Teknik SipilDocument4 pagesPoliteknik Negeri Medan: Laboratorium Teknik SipilBudi S F DamanikNo ratings yet
- Usle PDFDocument44 pagesUsle PDFtiaraNo ratings yet
- Planting Material Selection - FinDocument44 pagesPlanting Material Selection - FinApril Joy0% (2)
- Physiographic Division of IndiaDocument30 pagesPhysiographic Division of IndiaGurdarshan Singh Singh100% (1)
- 3rd Sem Report FDocument58 pages3rd Sem Report FpoojaNo ratings yet
- EFW 2203 Classification of Chirinda Forest Mapaure 680-1604-1-SMDocument9 pagesEFW 2203 Classification of Chirinda Forest Mapaure 680-1604-1-SMmuchy bNo ratings yet
- Cource OutlineDocument3 pagesCource OutlineabdulazizNo ratings yet
- Agricultural Superabsorbent PolymerDocument3 pagesAgricultural Superabsorbent PolymerjayalaheriNo ratings yet
- Wed Practice KeyDocument31 pagesWed Practice KeyRobyn Keil EdelsonNo ratings yet
- VL Dhan 207 and VL Dhan 208, The New High Yielding Varieties of Spring Rice For Uttarakhand HillsDocument4 pagesVL Dhan 207 and VL Dhan 208, The New High Yielding Varieties of Spring Rice For Uttarakhand HillsGrace CañasNo ratings yet
- Yellow River黃河 hs presentationDocument14 pagesYellow River黃河 hs presentationsky animal girlNo ratings yet