
Differential Pattern of Functional Brain Plasticity After Compassion and Empathy Training-Klimecki & Singer 2013
Differential Pattern of Functional Brain Plasticity After Compassion and Empathy Training-Klimecki & Singer 2013
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Icd 11 (2018)Document2,440 pagesIcd 11 (2018)Dr VJ George100% (2)
- Benedict SyndromeDocument5 pagesBenedict SyndromeMarthin Fernandes PasaribuNo ratings yet
- Unit 1 - Anatomy Study GuideDocument4 pagesUnit 1 - Anatomy Study Guide97baleNo ratings yet
- Mean Arterial PressureDocument3 pagesMean Arterial Pressureleslie vasquez lucumiNo ratings yet
- The Cns Portfolio EditionDocument9 pagesThe Cns Portfolio Editionapi-438492450No ratings yet
- Practice Short Answer Questions For REPRODUCTIVE PHYSIOLOGYDocument6 pagesPractice Short Answer Questions For REPRODUCTIVE PHYSIOLOGYKamNo ratings yet
- Hema - General Characteristics of Blood and Hematological TestsDocument5 pagesHema - General Characteristics of Blood and Hematological TestsBSMT Kharylle divine FuentibellaNo ratings yet
- Glossary of Greek and Latin Prefixes, Roots and SuffixesDocument6 pagesGlossary of Greek and Latin Prefixes, Roots and SuffixesDelia DeNo ratings yet
- Meditation Can Change Your BrainDocument38 pagesMeditation Can Change Your BrainZena Chames Eddin100% (1)
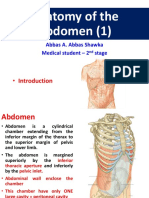
- Anatomy of The Abdomen (1) : - IntroductionDocument21 pagesAnatomy of The Abdomen (1) : - IntroductionMunachande KanondoNo ratings yet
- Activity 10 Circulatory System DiscussionDocument3 pagesActivity 10 Circulatory System DiscussionNathaniel FelicianoNo ratings yet
- Transfusion and Apheresis ScienceDocument6 pagesTransfusion and Apheresis Sciencemy accountNo ratings yet
- Science 6 2nd Quarter W3Document10 pagesScience 6 2nd Quarter W3Leo CerenoNo ratings yet
- Large Intestine - Structure and FunctionDocument13 pagesLarge Intestine - Structure and FunctiondivyasreekanthanNo ratings yet
- Demonstration of The Reflex Pathway in FrogsDocument12 pagesDemonstration of The Reflex Pathway in FrogsRuwiniRajakarunaNo ratings yet
- Human Reproduction - Part 1Document22 pagesHuman Reproduction - Part 1NickOoPandeyNo ratings yet
- Science Workbook (Bones and Muscles)Document83 pagesScience Workbook (Bones and Muscles)Reinhart LogronioNo ratings yet
- 2-Muscular System HandoutDocument4 pages2-Muscular System Handoutapi-302873465No ratings yet
- Introduction To HematologyDocument61 pagesIntroduction To HematologyThis is PonyNo ratings yet
- Activity 4 - Male Reproductive SystemDocument17 pagesActivity 4 - Male Reproductive Systemisabellamarie.castillo.crsNo ratings yet
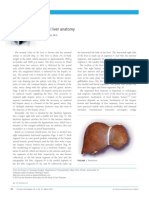
- Normal Liver AnatomyDocument3 pagesNormal Liver AnatomyZsolt Dubravcsik100% (1)
- Blood: Prepared by Patty Bostwick-Taylor, Florence-Darlington Technical CollegeDocument63 pagesBlood: Prepared by Patty Bostwick-Taylor, Florence-Darlington Technical Collegedomo domoNo ratings yet
- Age Changes in Enamel, Dentin and PulpDocument45 pagesAge Changes in Enamel, Dentin and Pulpanmol43% (7)
- The Power of Your Amazing BrainDocument3 pagesThe Power of Your Amazing BrainblackvenumNo ratings yet
- Oral Hygiene and OrthodonticsDocument17 pagesOral Hygiene and OrthodonticsDevangana DasNo ratings yet
- Closure of Midline Diastema by Direct Composite Resin Build-Up Using Putty Index - A CasereportDocument5 pagesClosure of Midline Diastema by Direct Composite Resin Build-Up Using Putty Index - A CasereportIJAR JOURNALNo ratings yet
- Genetic Aspect of Folliculogenesis: Budi WiwekoDocument72 pagesGenetic Aspect of Folliculogenesis: Budi WiwekoAgusNo ratings yet
- Daoh SyllabusDocument7 pagesDaoh SyllabusAsha SureshNo ratings yet
- A Multidisciplinary Treatment of A Dental Trauma Trauma DentaraDocument6 pagesA Multidisciplinary Treatment of A Dental Trauma Trauma DentaraStefana NanuNo ratings yet
- Zoology (T & E) Model Question PaperDocument8 pagesZoology (T & E) Model Question PaperPalanichamy MuthuNo ratings yet





































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Icd 11 (2018)Document2,440 pagesIcd 11 (2018)Dr VJ George100% (2)
- Benedict SyndromeDocument5 pagesBenedict SyndromeMarthin Fernandes PasaribuNo ratings yet
- Unit 1 - Anatomy Study GuideDocument4 pagesUnit 1 - Anatomy Study Guide97baleNo ratings yet
- Mean Arterial PressureDocument3 pagesMean Arterial Pressureleslie vasquez lucumiNo ratings yet
- The Cns Portfolio EditionDocument9 pagesThe Cns Portfolio Editionapi-438492450No ratings yet
- Practice Short Answer Questions For REPRODUCTIVE PHYSIOLOGYDocument6 pagesPractice Short Answer Questions For REPRODUCTIVE PHYSIOLOGYKamNo ratings yet
- Hema - General Characteristics of Blood and Hematological TestsDocument5 pagesHema - General Characteristics of Blood and Hematological TestsBSMT Kharylle divine FuentibellaNo ratings yet
- Glossary of Greek and Latin Prefixes, Roots and SuffixesDocument6 pagesGlossary of Greek and Latin Prefixes, Roots and SuffixesDelia DeNo ratings yet
- Meditation Can Change Your BrainDocument38 pagesMeditation Can Change Your BrainZena Chames Eddin100% (1)
- Anatomy of The Abdomen (1) : - IntroductionDocument21 pagesAnatomy of The Abdomen (1) : - IntroductionMunachande KanondoNo ratings yet
- Activity 10 Circulatory System DiscussionDocument3 pagesActivity 10 Circulatory System DiscussionNathaniel FelicianoNo ratings yet
- Transfusion and Apheresis ScienceDocument6 pagesTransfusion and Apheresis Sciencemy accountNo ratings yet
- Science 6 2nd Quarter W3Document10 pagesScience 6 2nd Quarter W3Leo CerenoNo ratings yet
- Large Intestine - Structure and FunctionDocument13 pagesLarge Intestine - Structure and FunctiondivyasreekanthanNo ratings yet
- Demonstration of The Reflex Pathway in FrogsDocument12 pagesDemonstration of The Reflex Pathway in FrogsRuwiniRajakarunaNo ratings yet
- Human Reproduction - Part 1Document22 pagesHuman Reproduction - Part 1NickOoPandeyNo ratings yet
- Science Workbook (Bones and Muscles)Document83 pagesScience Workbook (Bones and Muscles)Reinhart LogronioNo ratings yet
- 2-Muscular System HandoutDocument4 pages2-Muscular System Handoutapi-302873465No ratings yet
- Introduction To HematologyDocument61 pagesIntroduction To HematologyThis is PonyNo ratings yet
- Activity 4 - Male Reproductive SystemDocument17 pagesActivity 4 - Male Reproductive Systemisabellamarie.castillo.crsNo ratings yet
- Normal Liver AnatomyDocument3 pagesNormal Liver AnatomyZsolt Dubravcsik100% (1)
- Blood: Prepared by Patty Bostwick-Taylor, Florence-Darlington Technical CollegeDocument63 pagesBlood: Prepared by Patty Bostwick-Taylor, Florence-Darlington Technical Collegedomo domoNo ratings yet
- Age Changes in Enamel, Dentin and PulpDocument45 pagesAge Changes in Enamel, Dentin and Pulpanmol43% (7)
- The Power of Your Amazing BrainDocument3 pagesThe Power of Your Amazing BrainblackvenumNo ratings yet
- Oral Hygiene and OrthodonticsDocument17 pagesOral Hygiene and OrthodonticsDevangana DasNo ratings yet
- Closure of Midline Diastema by Direct Composite Resin Build-Up Using Putty Index - A CasereportDocument5 pagesClosure of Midline Diastema by Direct Composite Resin Build-Up Using Putty Index - A CasereportIJAR JOURNALNo ratings yet
- Genetic Aspect of Folliculogenesis: Budi WiwekoDocument72 pagesGenetic Aspect of Folliculogenesis: Budi WiwekoAgusNo ratings yet
- Daoh SyllabusDocument7 pagesDaoh SyllabusAsha SureshNo ratings yet
- A Multidisciplinary Treatment of A Dental Trauma Trauma DentaraDocument6 pagesA Multidisciplinary Treatment of A Dental Trauma Trauma DentaraStefana NanuNo ratings yet
- Zoology (T & E) Model Question PaperDocument8 pagesZoology (T & E) Model Question PaperPalanichamy MuthuNo ratings yet