
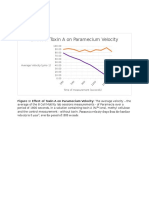
Cancer Malignancy Tumor
Cancer Malignancy Tumor
You might also like
- Biology 1A-Cell Motility Results and ConclusionsDocument6 pagesBiology 1A-Cell Motility Results and ConclusionsAna Maria100% (2)
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- B7-H3 Expression in Gastric CancerDocument6 pagesB7-H3 Expression in Gastric CancerIvoBarrosNo ratings yet
- Ournal of Linical Ncology: PurposeDocument7 pagesOurnal of Linical Ncology: PurposeIvor Wiguna Hartanto WilopoNo ratings yet
- Ni Hms 561057Document19 pagesNi Hms 561057Muhammad Reza IrzantoNo ratings yet
- Phase II Trial of Curcumin in Patients With Advanced Pancreatic Cancer.Document10 pagesPhase II Trial of Curcumin in Patients With Advanced Pancreatic Cancer.João FrancoNo ratings yet
- Decreased Histamine Catabolism in The Colonic Mucosa of Patients With Colonic AdenomaDocument7 pagesDecreased Histamine Catabolism in The Colonic Mucosa of Patients With Colonic AdenomaNejc KovačNo ratings yet
- Journal Pone 0031932Document9 pagesJournal Pone 0031932Joao FonsecaNo ratings yet
- SasdsadsaDocument11 pagesSasdsadsaJorge MariaNo ratings yet
- Cell Cycle Disorder and Gastrointestinal Cancers 2000Document8 pagesCell Cycle Disorder and Gastrointestinal Cancers 2000benmerzougNo ratings yet
- 2015 FBWX7 Not Prognosis in CRCDocument8 pages2015 FBWX7 Not Prognosis in CRCmaomaochongNo ratings yet
- Jurnal Prognosis 2Document8 pagesJurnal Prognosis 2Chlarissa WahabNo ratings yet
- Articulo de HistologiaDocument6 pagesArticulo de HistologiaalexbowwNo ratings yet
- Cam40002 0234Document9 pagesCam40002 0234Domenico LombardiniNo ratings yet
- Circulating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyDocument10 pagesCirculating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyftwNo ratings yet
- Plasma MesothelinDocument7 pagesPlasma Mesothelinpaulo saNo ratings yet
- 2018 Article 1232Document10 pages2018 Article 1232saveriomalara6No ratings yet
- Nejmoa 0908721Document9 pagesNejmoa 0908721Gustavo BraccoNo ratings yet
- Differential Expression of The Cck-A and Cck-B:gastrin Receptor Genes in Human Cancers of The Esophagus, Stomach and ColonDocument6 pagesDifferential Expression of The Cck-A and Cck-B:gastrin Receptor Genes in Human Cancers of The Esophagus, Stomach and ColonJaya Kunal DubeyNo ratings yet
- Cancer de PancreasDocument9 pagesCancer de PancreasferNo ratings yet
- ConutDocument9 pagesConutargamaviNo ratings yet
- 15-Li Chen (531-538)Document8 pages15-Li Chen (531-538)Nguyễn Thanh TùngNo ratings yet
- Ajcr0008 2436Document21 pagesAjcr0008 2436louisehip UFCNo ratings yet
- Palliative Chemotherapy For Advanced Colorectal Cancer Systemati 2000Document5 pagesPalliative Chemotherapy For Advanced Colorectal Cancer Systemati 2000Wiro SablengNo ratings yet
- Biomarkers For The Diagnosis of Cholangiocarcinoma A Systematic Review 2018Document10 pagesBiomarkers For The Diagnosis of Cholangiocarcinoma A Systematic Review 2018Anna MariaNo ratings yet
- Alterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasDocument11 pagesAlterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasAbdur Rachman Ba'abdullahNo ratings yet
- Sabcs 2014 AllabstractsDocument1,508 pagesSabcs 2014 Allabstractsrajesh4189No ratings yet
- Chromosomal Imbalances in Successive Moments of Human Bladder Urothelial CarcinomaDocument9 pagesChromosomal Imbalances in Successive Moments of Human Bladder Urothelial CarcinomayuenkeithNo ratings yet
- 1677 5538 Ibju 42 04 0694Document10 pages1677 5538 Ibju 42 04 0694Lucas VibiamNo ratings yet
- Christodouleas 2014Document9 pagesChristodouleas 2014nimaelhajjiNo ratings yet
- Antiproliferative Effects and Mechanisms of Liver X Receptor Ligands in Pancreatic Ductal Adenocarcinoma CellsDocument11 pagesAntiproliferative Effects and Mechanisms of Liver X Receptor Ligands in Pancreatic Ductal Adenocarcinoma CellsscribdenesimoNo ratings yet
- 10.1186@s13048 020 00619 6Document10 pages10.1186@s13048 020 00619 6Berry BancinNo ratings yet
- Jaaha Ms 6280Document8 pagesJaaha Ms 6280anna zanellaNo ratings yet
- Nej Me 1514353Document2 pagesNej Me 1514353anggiNo ratings yet
- CDDP Gemzar BilcanDocument9 pagesCDDP Gemzar BilcanhodimirekNo ratings yet
- International Seminars in Surgical OncologyDocument8 pagesInternational Seminars in Surgical OncologyAhmad ShafiqNo ratings yet
- Baumann 2013Document8 pagesBaumann 2013nimaelhajjiNo ratings yet
- Ahn2008 Article ClinicopathologicalFeaturesAndDocument8 pagesAhn2008 Article ClinicopathologicalFeaturesAndXavier QuinteroNo ratings yet
- Geneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. FieldsDocument26 pagesGeneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. Fieldsjomigp01No ratings yet
- Cancer de EsófagoDocument12 pagesCancer de EsófagoJuan Jose Sanchez HuamanNo ratings yet
- MSTF Guideline Bowel Cleansing Oct 2014Document18 pagesMSTF Guideline Bowel Cleansing Oct 2014CynthiaNo ratings yet
- Prognostic Factors in Nasopharyngeal Carcinoma With Synchronous Liver Metastasis: A Retrospective Study For The Management of TreatmentDocument7 pagesPrognostic Factors in Nasopharyngeal Carcinoma With Synchronous Liver Metastasis: A Retrospective Study For The Management of TreatmentChairul Nurdin AzaliNo ratings yet
- Impact of Tumor Location On Nodal Evaluation of Colon Cancer 12 Lymph Nodes 2008Document8 pagesImpact of Tumor Location On Nodal Evaluation of Colon Cancer 12 Lymph Nodes 2008MARIO DCARLO TREJO HUAMANNo ratings yet
- The OncologistDocument7 pagesThe OncologistManunart FeungpeanNo ratings yet
- Caner Gastrico en JovenesDocument13 pagesCaner Gastrico en JovenesandreaNo ratings yet
- Rugge 2018Document8 pagesRugge 2018Vanessa BecerraNo ratings yet
- Stem Loop PrimerDocument6 pagesStem Loop PrimerSyeda Farah ShahNo ratings yet
- Diagnostics 13 01362Document10 pagesDiagnostics 13 01362ela.sofiaNo ratings yet
- 172 - 04 101 13 PDFDocument8 pages172 - 04 101 13 PDFAlexandrosNo ratings yet
- Incidental Gallbladder Cancer. Residual Cancer Discovered at Oncologic Extended Resection Determines Outcome. A Report From High - and Low-IncidencDocument10 pagesIncidental Gallbladder Cancer. Residual Cancer Discovered at Oncologic Extended Resection Determines Outcome. A Report From High - and Low-IncidencCarlos SotoNo ratings yet
- Nej Mo A 0908721Document9 pagesNej Mo A 0908721Memble SangatNo ratings yet
- Microarreglos de Carcinoma Hepatocelular.Document14 pagesMicroarreglos de Carcinoma Hepatocelular.planhigion06No ratings yet
- Algoritma Kanker KolorektalDocument8 pagesAlgoritma Kanker KolorektalHasan NyambeNo ratings yet
- Prognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaDocument7 pagesPrognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaLuis FuentesNo ratings yet
- Expression of Survivin mRNA in Gallbladder Cancer - A Diagnostic and Prognostic Marker 1.29.38 PMDocument6 pagesExpression of Survivin mRNA in Gallbladder Cancer - A Diagnostic and Prognostic Marker 1.29.38 PMJaya Kunal DubeyNo ratings yet
- Thno 54961r2 3Document45 pagesThno 54961r2 3Eduardo JuarezNo ratings yet
- Colorectal CarcinomaDocument9 pagesColorectal CarcinomaKantona FrankNo ratings yet
- Role of Diet in The Management of Carcinoid Syndrome Clinical Recommendations For Nutrition in Patients With Neuroendocrine TumorsDocument11 pagesRole of Diet in The Management of Carcinoid Syndrome Clinical Recommendations For Nutrition in Patients With Neuroendocrine TumorsDoc Nelson JuniorNo ratings yet
- Paper 12Document7 pagesPaper 12shimaa youssifNo ratings yet
- AACR 2017 Proceedings: Abstracts 3063-5947From EverandAACR 2017 Proceedings: Abstracts 3063-5947No ratings yet
- MindMapper Tutorial PDFDocument1 pageMindMapper Tutorial PDFRidha Surya NugrahaNo ratings yet
- 2007 TAG SynapticDocument5 pages2007 TAG SynapticRidha Surya NugrahaNo ratings yet
- Bangs atDocument1 pageBangs atRidha Surya NugrahaNo ratings yet
- 2008 Synaptic Vesicle FusionDocument10 pages2008 Synaptic Vesicle FusionRidha Surya NugrahaNo ratings yet
- Matriks Kegiatan Blok 10Document2 pagesMatriks Kegiatan Blok 10Ridha Surya NugrahaNo ratings yet
- RAV Nonproliferative Diabetic Retinopathy NPDRDocument1 pageRAV Nonproliferative Diabetic Retinopathy NPDRRidha Surya NugrahaNo ratings yet
- Original Article: Clinical and Laboratory Features of Typhoid Fever in ChildhoodDocument6 pagesOriginal Article: Clinical and Laboratory Features of Typhoid Fever in ChildhoodRidha Surya NugrahaNo ratings yet
- Sdasldmk MCDMFL DSFDocument1 pageSdasldmk MCDMFL DSFRidha Surya NugrahaNo ratings yet
- (Transes) Human Histology - 10 Endocrine GlandsDocument10 pages(Transes) Human Histology - 10 Endocrine GlandsReina CastronuevoNo ratings yet
- 2962 Plasma Proteins PPT 53b668dea4a12Document35 pages2962 Plasma Proteins PPT 53b668dea4a12mahmoud fuqahaNo ratings yet
- Do Eosinophils Have A Role in The Severity of Babesia Annae Infection 2005 Veterinary ParasitologDocument2 pagesDo Eosinophils Have A Role in The Severity of Babesia Annae Infection 2005 Veterinary ParasitologGabriela Victoria MartinescuNo ratings yet
- Molecular Biology COVID-19 (SARS-Cov-2) RT-PCR: 80994699 Mr.I C SharmaDocument1 pageMolecular Biology COVID-19 (SARS-Cov-2) RT-PCR: 80994699 Mr.I C SharmaMohd YasarNo ratings yet
- International Biology Olympiad - Hong Kong Contest 2018 - 2018Document70 pagesInternational Biology Olympiad - Hong Kong Contest 2018 - 2018Ngọc CaiNo ratings yet
- Vargas, Bartolome Dr. Thornton, Karen 17675251Document1 pageVargas, Bartolome Dr. Thornton, Karen 17675251ahmedNo ratings yet
- DR Tahir Eye mcqs1Document10 pagesDR Tahir Eye mcqs1YUAN LINo ratings yet
- A Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientDocument6 pagesA Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientMichael Kivumbi100% (3)
- CV David C Eustice 1-2010Document6 pagesCV David C Eustice 1-2010Teresa MortonNo ratings yet
- Full Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions ManualDocument35 pagesFull Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions Manualudalbornhockeyspl2100% (31)
- 2020 AbsoluteDocument482 pages2020 AbsoluteAula AhmadNo ratings yet
- Respiratory System - WikipediaDocument1 pageRespiratory System - WikipediaMatias Kristel AnnNo ratings yet
- Patologi Tumor Sistim UrogenitalDocument29 pagesPatologi Tumor Sistim UrogenitalSiddiq Blackhell CakepNo ratings yet
- Premalignant LesionsDocument76 pagesPremalignant LesionsPrima D Andri100% (1)
- 1) Class 12th Biology Complete Unacademy BookDocument538 pages1) Class 12th Biology Complete Unacademy Bookkanikadudeja.07No ratings yet
- 1221 Adler Richard PDFDocument27 pages1221 Adler Richard PDFAvi BautistaNo ratings yet
- Bio 10 Master Study Guide Part 1Document9 pagesBio 10 Master Study Guide Part 1leonor.estimaNo ratings yet
- 21 382 ImmunologyDocument142 pages21 382 ImmunologyAmirabbas SaffariNo ratings yet
- Cellular AberrationDocument6 pagesCellular AberrationNeslie Lagare SamonteNo ratings yet
- Studi Kasus Bioetika Semester Gasal 2015-2016 PDFDocument8 pagesStudi Kasus Bioetika Semester Gasal 2015-2016 PDFShokhikhun NatiqNo ratings yet
- Small Animal OncologyDocument103 pagesSmall Animal Oncologyvet53No ratings yet
- DOT Passanten Versie Voor Communicatie 20211012Document1,566 pagesDOT Passanten Versie Voor Communicatie 20211012Dr RajeshNo ratings yet
- Female Pattern Hair Loss A Clinical and Pathophysiological PDFDocument15 pagesFemale Pattern Hair Loss A Clinical and Pathophysiological PDFJéssicaFigueredoSouzaNo ratings yet
- pRS415Document12 pagespRS415DiegoNo ratings yet
- Direct Comparison of Antimicrobial SusceptibilityDocument5 pagesDirect Comparison of Antimicrobial SusceptibilityRoela GeeNo ratings yet
- DisfungsiDocument1 pageDisfungsiHandayan HtbNo ratings yet
- What Is EpilepsyDocument5 pagesWhat Is Epilepsyhambysa12No ratings yet
- Glucose-6-Phosphate Dehydrogenase Deficiency - Overview: Symptom Treatment Prevention All InformationDocument2 pagesGlucose-6-Phosphate Dehydrogenase Deficiency - Overview: Symptom Treatment Prevention All InformationJinky TenezaNo ratings yet
- Case PresentationDocument21 pagesCase PresentationayhamsalimNo ratings yet
































































































Download as pdf or txt
You might also like
- Biology 1A-Cell Motility Results and ConclusionsDocument6 pagesBiology 1A-Cell Motility Results and ConclusionsAna Maria100% (2)
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- B7-H3 Expression in Gastric CancerDocument6 pagesB7-H3 Expression in Gastric CancerIvoBarrosNo ratings yet
- Ournal of Linical Ncology: PurposeDocument7 pagesOurnal of Linical Ncology: PurposeIvor Wiguna Hartanto WilopoNo ratings yet
- Ni Hms 561057Document19 pagesNi Hms 561057Muhammad Reza IrzantoNo ratings yet
- Phase II Trial of Curcumin in Patients With Advanced Pancreatic Cancer.Document10 pagesPhase II Trial of Curcumin in Patients With Advanced Pancreatic Cancer.João FrancoNo ratings yet
- Decreased Histamine Catabolism in The Colonic Mucosa of Patients With Colonic AdenomaDocument7 pagesDecreased Histamine Catabolism in The Colonic Mucosa of Patients With Colonic AdenomaNejc KovačNo ratings yet
- Journal Pone 0031932Document9 pagesJournal Pone 0031932Joao FonsecaNo ratings yet
- SasdsadsaDocument11 pagesSasdsadsaJorge MariaNo ratings yet
- Cell Cycle Disorder and Gastrointestinal Cancers 2000Document8 pagesCell Cycle Disorder and Gastrointestinal Cancers 2000benmerzougNo ratings yet
- 2015 FBWX7 Not Prognosis in CRCDocument8 pages2015 FBWX7 Not Prognosis in CRCmaomaochongNo ratings yet
- Jurnal Prognosis 2Document8 pagesJurnal Prognosis 2Chlarissa WahabNo ratings yet
- Articulo de HistologiaDocument6 pagesArticulo de HistologiaalexbowwNo ratings yet
- Cam40002 0234Document9 pagesCam40002 0234Domenico LombardiniNo ratings yet
- Circulating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyDocument10 pagesCirculating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyftwNo ratings yet
- Plasma MesothelinDocument7 pagesPlasma Mesothelinpaulo saNo ratings yet
- 2018 Article 1232Document10 pages2018 Article 1232saveriomalara6No ratings yet
- Nejmoa 0908721Document9 pagesNejmoa 0908721Gustavo BraccoNo ratings yet
- Differential Expression of The Cck-A and Cck-B:gastrin Receptor Genes in Human Cancers of The Esophagus, Stomach and ColonDocument6 pagesDifferential Expression of The Cck-A and Cck-B:gastrin Receptor Genes in Human Cancers of The Esophagus, Stomach and ColonJaya Kunal DubeyNo ratings yet
- Cancer de PancreasDocument9 pagesCancer de PancreasferNo ratings yet
- ConutDocument9 pagesConutargamaviNo ratings yet
- 15-Li Chen (531-538)Document8 pages15-Li Chen (531-538)Nguyễn Thanh TùngNo ratings yet
- Ajcr0008 2436Document21 pagesAjcr0008 2436louisehip UFCNo ratings yet
- Palliative Chemotherapy For Advanced Colorectal Cancer Systemati 2000Document5 pagesPalliative Chemotherapy For Advanced Colorectal Cancer Systemati 2000Wiro SablengNo ratings yet
- Biomarkers For The Diagnosis of Cholangiocarcinoma A Systematic Review 2018Document10 pagesBiomarkers For The Diagnosis of Cholangiocarcinoma A Systematic Review 2018Anna MariaNo ratings yet
- Alterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasDocument11 pagesAlterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasAbdur Rachman Ba'abdullahNo ratings yet
- Sabcs 2014 AllabstractsDocument1,508 pagesSabcs 2014 Allabstractsrajesh4189No ratings yet
- Chromosomal Imbalances in Successive Moments of Human Bladder Urothelial CarcinomaDocument9 pagesChromosomal Imbalances in Successive Moments of Human Bladder Urothelial CarcinomayuenkeithNo ratings yet
- 1677 5538 Ibju 42 04 0694Document10 pages1677 5538 Ibju 42 04 0694Lucas VibiamNo ratings yet
- Christodouleas 2014Document9 pagesChristodouleas 2014nimaelhajjiNo ratings yet
- Antiproliferative Effects and Mechanisms of Liver X Receptor Ligands in Pancreatic Ductal Adenocarcinoma CellsDocument11 pagesAntiproliferative Effects and Mechanisms of Liver X Receptor Ligands in Pancreatic Ductal Adenocarcinoma CellsscribdenesimoNo ratings yet
- 10.1186@s13048 020 00619 6Document10 pages10.1186@s13048 020 00619 6Berry BancinNo ratings yet
- Jaaha Ms 6280Document8 pagesJaaha Ms 6280anna zanellaNo ratings yet
- Nej Me 1514353Document2 pagesNej Me 1514353anggiNo ratings yet
- CDDP Gemzar BilcanDocument9 pagesCDDP Gemzar BilcanhodimirekNo ratings yet
- International Seminars in Surgical OncologyDocument8 pagesInternational Seminars in Surgical OncologyAhmad ShafiqNo ratings yet
- Baumann 2013Document8 pagesBaumann 2013nimaelhajjiNo ratings yet
- Ahn2008 Article ClinicopathologicalFeaturesAndDocument8 pagesAhn2008 Article ClinicopathologicalFeaturesAndXavier QuinteroNo ratings yet
- Geneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. FieldsDocument26 pagesGeneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. Fieldsjomigp01No ratings yet
- Cancer de EsófagoDocument12 pagesCancer de EsófagoJuan Jose Sanchez HuamanNo ratings yet
- MSTF Guideline Bowel Cleansing Oct 2014Document18 pagesMSTF Guideline Bowel Cleansing Oct 2014CynthiaNo ratings yet
- Prognostic Factors in Nasopharyngeal Carcinoma With Synchronous Liver Metastasis: A Retrospective Study For The Management of TreatmentDocument7 pagesPrognostic Factors in Nasopharyngeal Carcinoma With Synchronous Liver Metastasis: A Retrospective Study For The Management of TreatmentChairul Nurdin AzaliNo ratings yet
- Impact of Tumor Location On Nodal Evaluation of Colon Cancer 12 Lymph Nodes 2008Document8 pagesImpact of Tumor Location On Nodal Evaluation of Colon Cancer 12 Lymph Nodes 2008MARIO DCARLO TREJO HUAMANNo ratings yet
- The OncologistDocument7 pagesThe OncologistManunart FeungpeanNo ratings yet
- Caner Gastrico en JovenesDocument13 pagesCaner Gastrico en JovenesandreaNo ratings yet
- Rugge 2018Document8 pagesRugge 2018Vanessa BecerraNo ratings yet
- Stem Loop PrimerDocument6 pagesStem Loop PrimerSyeda Farah ShahNo ratings yet
- Diagnostics 13 01362Document10 pagesDiagnostics 13 01362ela.sofiaNo ratings yet
- 172 - 04 101 13 PDFDocument8 pages172 - 04 101 13 PDFAlexandrosNo ratings yet
- Incidental Gallbladder Cancer. Residual Cancer Discovered at Oncologic Extended Resection Determines Outcome. A Report From High - and Low-IncidencDocument10 pagesIncidental Gallbladder Cancer. Residual Cancer Discovered at Oncologic Extended Resection Determines Outcome. A Report From High - and Low-IncidencCarlos SotoNo ratings yet
- Nej Mo A 0908721Document9 pagesNej Mo A 0908721Memble SangatNo ratings yet
- Microarreglos de Carcinoma Hepatocelular.Document14 pagesMicroarreglos de Carcinoma Hepatocelular.planhigion06No ratings yet
- Algoritma Kanker KolorektalDocument8 pagesAlgoritma Kanker KolorektalHasan NyambeNo ratings yet
- Prognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaDocument7 pagesPrognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaLuis FuentesNo ratings yet
- Expression of Survivin mRNA in Gallbladder Cancer - A Diagnostic and Prognostic Marker 1.29.38 PMDocument6 pagesExpression of Survivin mRNA in Gallbladder Cancer - A Diagnostic and Prognostic Marker 1.29.38 PMJaya Kunal DubeyNo ratings yet
- Thno 54961r2 3Document45 pagesThno 54961r2 3Eduardo JuarezNo ratings yet
- Colorectal CarcinomaDocument9 pagesColorectal CarcinomaKantona FrankNo ratings yet
- Role of Diet in The Management of Carcinoid Syndrome Clinical Recommendations For Nutrition in Patients With Neuroendocrine TumorsDocument11 pagesRole of Diet in The Management of Carcinoid Syndrome Clinical Recommendations For Nutrition in Patients With Neuroendocrine TumorsDoc Nelson JuniorNo ratings yet
- Paper 12Document7 pagesPaper 12shimaa youssifNo ratings yet
- AACR 2017 Proceedings: Abstracts 3063-5947From EverandAACR 2017 Proceedings: Abstracts 3063-5947No ratings yet
- MindMapper Tutorial PDFDocument1 pageMindMapper Tutorial PDFRidha Surya NugrahaNo ratings yet
- 2007 TAG SynapticDocument5 pages2007 TAG SynapticRidha Surya NugrahaNo ratings yet
- Bangs atDocument1 pageBangs atRidha Surya NugrahaNo ratings yet
- 2008 Synaptic Vesicle FusionDocument10 pages2008 Synaptic Vesicle FusionRidha Surya NugrahaNo ratings yet
- Matriks Kegiatan Blok 10Document2 pagesMatriks Kegiatan Blok 10Ridha Surya NugrahaNo ratings yet
- RAV Nonproliferative Diabetic Retinopathy NPDRDocument1 pageRAV Nonproliferative Diabetic Retinopathy NPDRRidha Surya NugrahaNo ratings yet
- Original Article: Clinical and Laboratory Features of Typhoid Fever in ChildhoodDocument6 pagesOriginal Article: Clinical and Laboratory Features of Typhoid Fever in ChildhoodRidha Surya NugrahaNo ratings yet
- Sdasldmk MCDMFL DSFDocument1 pageSdasldmk MCDMFL DSFRidha Surya NugrahaNo ratings yet
- (Transes) Human Histology - 10 Endocrine GlandsDocument10 pages(Transes) Human Histology - 10 Endocrine GlandsReina CastronuevoNo ratings yet
- 2962 Plasma Proteins PPT 53b668dea4a12Document35 pages2962 Plasma Proteins PPT 53b668dea4a12mahmoud fuqahaNo ratings yet
- Do Eosinophils Have A Role in The Severity of Babesia Annae Infection 2005 Veterinary ParasitologDocument2 pagesDo Eosinophils Have A Role in The Severity of Babesia Annae Infection 2005 Veterinary ParasitologGabriela Victoria MartinescuNo ratings yet
- Molecular Biology COVID-19 (SARS-Cov-2) RT-PCR: 80994699 Mr.I C SharmaDocument1 pageMolecular Biology COVID-19 (SARS-Cov-2) RT-PCR: 80994699 Mr.I C SharmaMohd YasarNo ratings yet
- International Biology Olympiad - Hong Kong Contest 2018 - 2018Document70 pagesInternational Biology Olympiad - Hong Kong Contest 2018 - 2018Ngọc CaiNo ratings yet
- Vargas, Bartolome Dr. Thornton, Karen 17675251Document1 pageVargas, Bartolome Dr. Thornton, Karen 17675251ahmedNo ratings yet
- DR Tahir Eye mcqs1Document10 pagesDR Tahir Eye mcqs1YUAN LINo ratings yet
- A Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientDocument6 pagesA Project Proposal On Case Study and Management of A Type 1 Diabetes Mellitus ClientMichael Kivumbi100% (3)
- CV David C Eustice 1-2010Document6 pagesCV David C Eustice 1-2010Teresa MortonNo ratings yet
- Full Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions ManualDocument35 pagesFull Download Essentials of Physical Anthropology 10th Edition Jurmain Solutions Manualudalbornhockeyspl2100% (31)
- 2020 AbsoluteDocument482 pages2020 AbsoluteAula AhmadNo ratings yet
- Respiratory System - WikipediaDocument1 pageRespiratory System - WikipediaMatias Kristel AnnNo ratings yet
- Patologi Tumor Sistim UrogenitalDocument29 pagesPatologi Tumor Sistim UrogenitalSiddiq Blackhell CakepNo ratings yet
- Premalignant LesionsDocument76 pagesPremalignant LesionsPrima D Andri100% (1)
- 1) Class 12th Biology Complete Unacademy BookDocument538 pages1) Class 12th Biology Complete Unacademy Bookkanikadudeja.07No ratings yet
- 1221 Adler Richard PDFDocument27 pages1221 Adler Richard PDFAvi BautistaNo ratings yet
- Bio 10 Master Study Guide Part 1Document9 pagesBio 10 Master Study Guide Part 1leonor.estimaNo ratings yet
- 21 382 ImmunologyDocument142 pages21 382 ImmunologyAmirabbas SaffariNo ratings yet
- Cellular AberrationDocument6 pagesCellular AberrationNeslie Lagare SamonteNo ratings yet
- Studi Kasus Bioetika Semester Gasal 2015-2016 PDFDocument8 pagesStudi Kasus Bioetika Semester Gasal 2015-2016 PDFShokhikhun NatiqNo ratings yet
- Small Animal OncologyDocument103 pagesSmall Animal Oncologyvet53No ratings yet
- DOT Passanten Versie Voor Communicatie 20211012Document1,566 pagesDOT Passanten Versie Voor Communicatie 20211012Dr RajeshNo ratings yet
- Female Pattern Hair Loss A Clinical and Pathophysiological PDFDocument15 pagesFemale Pattern Hair Loss A Clinical and Pathophysiological PDFJéssicaFigueredoSouzaNo ratings yet
- pRS415Document12 pagespRS415DiegoNo ratings yet
- Direct Comparison of Antimicrobial SusceptibilityDocument5 pagesDirect Comparison of Antimicrobial SusceptibilityRoela GeeNo ratings yet
- DisfungsiDocument1 pageDisfungsiHandayan HtbNo ratings yet
- What Is EpilepsyDocument5 pagesWhat Is Epilepsyhambysa12No ratings yet
- Glucose-6-Phosphate Dehydrogenase Deficiency - Overview: Symptom Treatment Prevention All InformationDocument2 pagesGlucose-6-Phosphate Dehydrogenase Deficiency - Overview: Symptom Treatment Prevention All InformationJinky TenezaNo ratings yet
- Case PresentationDocument21 pagesCase PresentationayhamsalimNo ratings yet