Download as pdf or txt
You might also like
- Max Lab ReportDocument1 pageMax Lab ReportKallu PrasadNo ratings yet
- Hem Lab Microhematocrit F15Document10 pagesHem Lab Microhematocrit F15Joan Christie WijayaNo ratings yet
- 1862ec-1877ec-1867ec 2021-11 PDFDocument53 pages1862ec-1877ec-1867ec 2021-11 PDFJose Adán Abdala PalmaNo ratings yet
- PPS Core PediatricsDocument53 pagesPPS Core PediatricsDenise Castro100% (3)
- Immunohematology Trans by KTRC (Wala Ito Sa Book)Document21 pagesImmunohematology Trans by KTRC (Wala Ito Sa Book)Angelo ErispeNo ratings yet
- Supplemental DataRevisedDocument12 pagesSupplemental DataRevisedCezary TomczykNo ratings yet
- Cancer-Associated Adipocytes Promote The Invasion and Metastasis in Breast Cancer Through LIFCXCLs Positive Feedback Loop Ayesha MudassarDocument35 pagesCancer-Associated Adipocytes Promote The Invasion and Metastasis in Breast Cancer Through LIFCXCLs Positive Feedback Loop Ayesha MudassarAyesha MudassarNo ratings yet
- Ecm210 ProtocolDocument8 pagesEcm210 ProtocolaidanaNo ratings yet
- Cells Preferentially Vp3 And/or: T in Psoriatic Lesions Use T-Cell Receptor GenesDocument15 pagesCells Preferentially Vp3 And/or: T in Psoriatic Lesions Use T-Cell Receptor GenesSnehal BhagatNo ratings yet
- 2960 FullDocument7 pages2960 FullNela SharonNo ratings yet
- 靶向CD138的第四代CAR-T细胞的构建及功能研究 刘宇Document10 pages靶向CD138的第四代CAR-T细胞的构建及功能研究 刘宇Tianliang GuoNo ratings yet
- Supplemetary MaterialDocument5 pagesSupplemetary MaterialAnaNo ratings yet
- Bio Protocol689 SpleenDocument4 pagesBio Protocol689 SpleenBagus Muhammad IhsanNo ratings yet
- Toxoplasmose Latente EsplenomegaliaDocument11 pagesToxoplasmose Latente EsplenomegaliaWeverson LinharesNo ratings yet
- #Material Suplementar - Ref-56 Do Artigo - 2019-Metal Complexes of Flavonoids...Document7 pages#Material Suplementar - Ref-56 Do Artigo - 2019-Metal Complexes of Flavonoids...las.chemicalNo ratings yet
- Antibody Binding Profile of Purified and Cell-Bound CD26 1993Document14 pagesAntibody Binding Profile of Purified and Cell-Bound CD26 1993heyligenNo ratings yet
- Nanooptogenetic CART Cell ImmunotherapyMethods in Molecular Biology Clifton NJDocument12 pagesNanooptogenetic CART Cell ImmunotherapyMethods in Molecular Biology Clifton NJAlexNo ratings yet
- Laboratory Procedure Manual: Cd4+ T-Lymphocytes and Cd8 T Cells Whole Blood Tricount MethodDocument11 pagesLaboratory Procedure Manual: Cd4+ T-Lymphocytes and Cd8 T Cells Whole Blood Tricount MethodanggaririnNo ratings yet
- Methyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin CytoskeletonDocument6 pagesMethyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin CytoskeletonGiggly HadidNo ratings yet
- Candida 1Document10 pagesCandida 1mufqifitra160491No ratings yet
- ParasitDocument7 pagesParasitRainz BlackNo ratings yet
- Practical of Immunology (Lab-3)Document22 pagesPractical of Immunology (Lab-3)azeema fatimaNo ratings yet
- Ex Vivo T Cell Activation Assay - rCAPDocument3 pagesEx Vivo T Cell Activation Assay - rCAPMARIO RAMIREZ POZONo ratings yet
- MGCTDocument11 pagesMGCTcandiddreamsNo ratings yet
- His 12031Document4 pagesHis 12031JeevikaGoyalNo ratings yet
- Citotoxic CAPEDocument8 pagesCitotoxic CAPEannisa_yuniartiNo ratings yet
- Detection and Identification of Mycobacterium Species by Polymerase Chain Reaction (PCR) From Paraffin-Embedded Tissue Compare To AFB Staining in Pathological SectionsDocument6 pagesDetection and Identification of Mycobacterium Species by Polymerase Chain Reaction (PCR) From Paraffin-Embedded Tissue Compare To AFB Staining in Pathological Sectionsธิรดา สายสตรอง สายจำปาNo ratings yet
- Research Article: The c-MYC Protooncogene Expression in CholesteatomaDocument7 pagesResearch Article: The c-MYC Protooncogene Expression in CholesteatomaVicky G OeiNo ratings yet
- Genotoxicity StudyDocument6 pagesGenotoxicity StudyShraddha RautNo ratings yet
- Suppression of ClC-3 Channel Expression Reduces Migration of Nasopharyngeal Carcinoma CellsDocument11 pagesSuppression of ClC-3 Channel Expression Reduces Migration of Nasopharyngeal Carcinoma Cellsa0965828895No ratings yet
- Immuno-Monitoring of CD8+ T Cells in Whole Blood Versus PBMC SamplesDocument8 pagesImmuno-Monitoring of CD8+ T Cells in Whole Blood Versus PBMC SamplesastheniiaNo ratings yet
- A Simplified Method of Three Dimensional Technique For The Detection of AmpC Beta-LactamasesDocument7 pagesA Simplified Method of Three Dimensional Technique For The Detection of AmpC Beta-LactamasesInternational Medical PublisherNo ratings yet
- Immunol 1995 Kennedy 203 9Document7 pagesImmunol 1995 Kennedy 203 9Sholeh RhazesNo ratings yet
- Immunology00120 0028Document7 pagesImmunology00120 0028carlitoseverectNo ratings yet
- Lugtenberg 75 ElectrophoreticDocument5 pagesLugtenberg 75 ElectrophoreticYsmael E. HernandezNo ratings yet
- 0903197106siDocument8 pages0903197106siprism1702No ratings yet
- Cells and ReagentsDocument4 pagesCells and ReagentsSiva AllaNo ratings yet
- Erum Ahmed - MBP Final ReportDocument19 pagesErum Ahmed - MBP Final ReportShana FullerNo ratings yet
- Letters: Isolation of Rare Circulating Tumour Cells in Cancer Patients by Microchip TechnologyDocument8 pagesLetters: Isolation of Rare Circulating Tumour Cells in Cancer Patients by Microchip TechnologyFebrina ViselitaNo ratings yet
- Marrow Transplanted Patients Inhibits Cell-Mediated Cytolysis A Soluble Factor Released by CD8+CD57+ Lymphocytes From BoneDocument6 pagesMarrow Transplanted Patients Inhibits Cell-Mediated Cytolysis A Soluble Factor Released by CD8+CD57+ Lymphocytes From Bonecyahayue5554No ratings yet
- Cash Saving Recommendations For Heterocyclic CompoundsDocument2 pagesCash Saving Recommendations For Heterocyclic Compoundsokrabrown85No ratings yet
- Allogeneic Expanded Human Peripheral NK Cells Control Prostate Cancer Growth in A Preclinical Mouse Model of Castration-Resistant Prostate CancerDocument11 pagesAllogeneic Expanded Human Peripheral NK Cells Control Prostate Cancer Growth in A Preclinical Mouse Model of Castration-Resistant Prostate Cancerhoangphuong08101992No ratings yet
- C C CC CDocument18 pagesC C CC CbiopeteNo ratings yet
- Mto 201615Document7 pagesMto 201615Tim TiemannNo ratings yet
- Isolation of Stem Cells From Human Pancreatic CancerDocument4 pagesIsolation of Stem Cells From Human Pancreatic CancerAntonio mlnNo ratings yet
- Cisplatin Cell Cicle ArrestDocument7 pagesCisplatin Cell Cicle ArrestGabriele KrauseNo ratings yet
- Sample Lab ReportDocument5 pagesSample Lab Reportapi-232072092No ratings yet
- Cell CountDocument2 pagesCell CountBridget HaleyNo ratings yet
- Effects of Sodium Ascorbate (Vitamin C) and 2-Methyl-1,4-Naphthoquinone (Vitamin K3) Treatment On Human Tumor Cell Growth in VitroDocument6 pagesEffects of Sodium Ascorbate (Vitamin C) and 2-Methyl-1,4-Naphthoquinone (Vitamin K3) Treatment On Human Tumor Cell Growth in VitroUtopiaawaits100% (1)
- 1 s2.0 S0006497121069597 MainDocument13 pages1 s2.0 S0006497121069597 Mainf2xt4fj858No ratings yet
- 5689 FullDocument8 pages5689 FullAshish SharmaNo ratings yet
- Collective Lab Report 2 MolecularDocument12 pagesCollective Lab Report 2 Molecularmariam farragNo ratings yet
- 1-Eastern Mediterranean Health JournalDocument5 pages1-Eastern Mediterranean Health Journalreadyboy89No ratings yet
- Efectos Inmunoterapeuticos en Cancer de CPG en Modelo de Tumor Murino HYEON CHEOL CHO 2008Document7 pagesEfectos Inmunoterapeuticos en Cancer de CPG en Modelo de Tumor Murino HYEON CHEOL CHO 2008Ramiro J. Rodriguez GarciaNo ratings yet
- Designation:: Homo Sapiens (Human)Document2 pagesDesignation:: Homo Sapiens (Human)joaquimNo ratings yet
- Integrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell ReprogrammingDocument16 pagesIntegrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell Reprogrammingniccolo.liorni1No ratings yet
- Edwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Document48 pagesEdwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Edwin DarmawanNo ratings yet
- Data SupplementDocument17 pagesData SupplementManovriti ThakurNo ratings yet
- Method: Prevalence and molecular characterization of extended-spectrum β-lactamase-producing Klebsiella pneumoniaeDocument4 pagesMethod: Prevalence and molecular characterization of extended-spectrum β-lactamase-producing Klebsiella pneumoniaeAnupa PokhrelNo ratings yet
- tmpD44A TMPDocument15 pagestmpD44A TMPFrontiersNo ratings yet
- 1099 Full PDFDocument6 pages1099 Full PDFWahyu WijayantoNo ratings yet
- Clinical Manufacturing of CAR T Cells FoundationDocument7 pagesClinical Manufacturing of CAR T Cells FoundationMinh Trần ThịNo ratings yet
- MMP10 NSCLCDocument9 pagesMMP10 NSCLCbhaskarNo ratings yet
- Certificate For COVID-19 Vaccination: Beneficiary DetailsDocument1 pageCertificate For COVID-19 Vaccination: Beneficiary DetailsRavindra ErabattiNo ratings yet
- EUA AbtDX BNAgselftest IfuDocument20 pagesEUA AbtDX BNAgselftest IfuLUIS ALEJANDRO MURGASNo ratings yet
- Mast Cell Chymase Protects Against Acute Ischemic Kidney Injury by Limiting Neutrophil Hyperactivation and RecruitmentDocument40 pagesMast Cell Chymase Protects Against Acute Ischemic Kidney Injury by Limiting Neutrophil Hyperactivation and RecruitmentOmar Arias AndradeNo ratings yet
- Septic ArthritisDocument8 pagesSeptic ArthritisLorebell100% (2)
- Live Attenuated RSVvaccines CollinsDocument9 pagesLive Attenuated RSVvaccines CollinsYan MartinezNo ratings yet
- Rle BSN 2 (Lesson 3) (2021-2022)Document26 pagesRle BSN 2 (Lesson 3) (2021-2022)Flonamae AbiarNo ratings yet
- How To Protect Yourself and Others: Know How It SpreadsDocument2 pagesHow To Protect Yourself and Others: Know How It SpreadsNEWS CENTER Maine100% (1)
- Orchitis 160406115741Document28 pagesOrchitis 160406115741alyne100% (1)
- HIF-Mediated Erythropoiesis: HIF Overcomes Inflammatory InterferenceDocument1 pageHIF-Mediated Erythropoiesis: HIF Overcomes Inflammatory InterferenceLyra LorcaNo ratings yet
- AzathioprineDocument3 pagesAzathioprinesky.blueNo ratings yet
- KP White Blood Cell DisordersDocument15 pagesKP White Blood Cell DisorderssaskiakonitaNo ratings yet
- Systemic Lupus Erythematosus SLE - A Review of CliDocument5 pagesSystemic Lupus Erythematosus SLE - A Review of CliJafrinta Irma R.A.No ratings yet
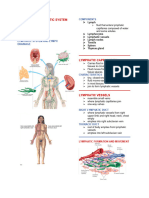
- Chapter 14 - Lymphatic SystemDocument10 pagesChapter 14 - Lymphatic SystemmargaretNo ratings yet
- Virology: EnterovirusesDocument40 pagesVirology: EnterovirusesMeevie ToledoNo ratings yet
- Mamalia Indonesia Diversitas Dan Potensi Sebagai Reservoir Penyakit ZoonosisDocument19 pagesMamalia Indonesia Diversitas Dan Potensi Sebagai Reservoir Penyakit ZoonosisYoga Sahria SimoNo ratings yet
- Erythematous Candidiasis PDFDocument2 pagesErythematous Candidiasis PDFPatrickNo ratings yet
- SARS-CoV-2 Viremia Is Associated With Distinct Proteomic Pathways and Predicts COVID-19 OutcomesDocument13 pagesSARS-CoV-2 Viremia Is Associated With Distinct Proteomic Pathways and Predicts COVID-19 OutcomesYoga Nugraha WNo ratings yet
- Psoriatic Arthritis - StatPearls - NCBI BookshelfDocument14 pagesPsoriatic Arthritis - StatPearls - NCBI BookshelfDhany karubuyNo ratings yet
- Ananthanarayan & Paniker's Textbook of Microbiology, 2005Document672 pagesAnanthanarayan & Paniker's Textbook of Microbiology, 2005Naga Raju Kandula67% (3)
- "Dengue and Dengue Hemorrhagic Fever": Angeles University FoundationDocument17 pages"Dengue and Dengue Hemorrhagic Fever": Angeles University FoundationLois DanielleNo ratings yet
- Jurnal Integumen, Ahmad Najib 2107028Document9 pagesJurnal Integumen, Ahmad Najib 2107028Ahmad NajibNo ratings yet
- AbstrakDocument7 pagesAbstrakAriandindi AriandiNo ratings yet
- ENTEROBACTERIACEAEDocument11 pagesENTEROBACTERIACEAEYormae QuezonNo ratings yet
- COVIDVaccineResponse 22dec2021 FINALDocument2 pagesCOVIDVaccineResponse 22dec2021 FINALTara Copp100% (1)
- Haemophilus DucreyiDocument15 pagesHaemophilus DucreyichristieNo ratings yet
- Empirical Prescribing Chart GlasgowDocument1 pageEmpirical Prescribing Chart GlasgowHariharan Narendran75% (4)