
Journal Lbm4
Journal Lbm4
You might also like
- High Yield NBME Images AnnotatedDocument84 pagesHigh Yield NBME Images AnnotatedNareman Abd El HadeNo ratings yet
- Diagnosis of Small Lung Biopsy An Integrated Approach by Mostafa M. FraigDocument111 pagesDiagnosis of Small Lung Biopsy An Integrated Approach by Mostafa M. FraigFlaviaNo ratings yet
- (Medical Masterclass) Coll.-Infectious Diseases and DermatologyDocument333 pages(Medical Masterclass) Coll.-Infectious Diseases and DermatologyVijay Mg100% (2)
- Microscopic Slides Pathology-2Document135 pagesMicroscopic Slides Pathology-2Justin JannatiNo ratings yet
- Immune Response To SchistosomaDocument11 pagesImmune Response To SchistosomaRobMarvinNo ratings yet
- Blastocytosis Hominis - The Summary From A To ZDocument11 pagesBlastocytosis Hominis - The Summary From A To ZSalsabila TasyaNo ratings yet
- Schistosoma "Eggs-Iting" The Host: Granuloma Formation and Egg ExcretionDocument16 pagesSchistosoma "Eggs-Iting" The Host: Granuloma Formation and Egg ExcretionRoberto Evangelista CostaNo ratings yet
- Schistosomiasis and Liver Fibrosis: Review ArticleDocument8 pagesSchistosomiasis and Liver Fibrosis: Review ArticlevigneshNo ratings yet
- Jordan University Faculty of Dentistry DR Mohammad Al-Tamimi, MD, PHDDocument20 pagesJordan University Faculty of Dentistry DR Mohammad Al-Tamimi, MD, PHDDaniel AtiehNo ratings yet
- PatogenesisDocument28 pagesPatogenesisOka WidyaningsihNo ratings yet
- Fimmu 04 00507 PDFDocument20 pagesFimmu 04 00507 PDFSilma FarrahaNo ratings yet
- BMJ d2651 PDFDocument11 pagesBMJ d2651 PDFKarin KurniatiNo ratings yet
- Key Infections in The PlacentaDocument14 pagesKey Infections in The PlacentaLauraGonzalezNo ratings yet
- Schistosomiasis: ArticleDocument20 pagesSchistosomiasis: ArticleVincent ReyesNo ratings yet
- Salmonella, The Host and Disease: A Brief ReviewDocument8 pagesSalmonella, The Host and Disease: A Brief ReviewPedro Albán MNo ratings yet
- 2015 Kline Bowdish Infections in An Aging PopulationDocument5 pages2015 Kline Bowdish Infections in An Aging PopulationLydiaphNo ratings yet
- Jiu 189Document9 pagesJiu 189barreirosanchez984No ratings yet
- Petton 2021 Frontiers Pacific MANUSCRIT2Document26 pagesPetton 2021 Frontiers Pacific MANUSCRIT2AmadorRevillaNo ratings yet
- Gram-Positive Cocci Staphylococcus CLINICAL SIGNIFICANCEDocument8 pagesGram-Positive Cocci Staphylococcus CLINICAL SIGNIFICANCEsobia faryadaliNo ratings yet
- Advances in New Target Molecules Against - Peng ZhuDocument22 pagesAdvances in New Target Molecules Against - Peng Zhuana.pinho.souzaNo ratings yet
- Human SchistosomiasisDocument18 pagesHuman SchistosomiasisbassbngNo ratings yet
- Othman 2014Document44 pagesOthman 2014Jessica QuevedoNo ratings yet
- 6 Helminth2 - PPTDocument8 pages6 Helminth2 - PPTRaihan Zulfiqar HalimNo ratings yet
- Arch Dis Child 1998 Ladhani 85 8Document5 pagesArch Dis Child 1998 Ladhani 85 8Dian Ayu Permata SandiNo ratings yet
- Blastocystis How Do Specific Diets and Human Gut MicrobiotaDocument10 pagesBlastocystis How Do Specific Diets and Human Gut MicrobiotaVanessa OcegueraNo ratings yet
- Empyema 2Document9 pagesEmpyema 2phobicmdNo ratings yet
- Lyon 2005Document21 pagesLyon 2005Renata StefaniNo ratings yet
- At The Turn of The 20th CenturyDocument6 pagesAt The Turn of The 20th CenturyStefanosSokratousNo ratings yet
- Location and Pathogenic Potenctial of Blastocystis in The Porcine IntestineDocument8 pagesLocation and Pathogenic Potenctial of Blastocystis in The Porcine Intestinesebasrd07No ratings yet
- Major Serological Screening Methodologies To Anti - T. Gondii Antibodies Detection in Pregnancy: Mini Literature ViewDocument4 pagesMajor Serological Screening Methodologies To Anti - T. Gondii Antibodies Detection in Pregnancy: Mini Literature Viewpublishing groupNo ratings yet
- Pancreatic Tuberculosis Review ArticleDocument24 pagesPancreatic Tuberculosis Review ArticlesamudraandiNo ratings yet
- Isospora SuisDocument35 pagesIsospora SuisAfif DihanNo ratings yet
- Interactions Between Parasites and Human MicrobiotDocument6 pagesInteractions Between Parasites and Human MicrobiotGregorio De Las CasasNo ratings yet
- Toxo PregnancyDocument15 pagesToxo PregnancyYossy CatarinaNo ratings yet
- First Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDocument11 pagesFirst Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDavid A. Osorio G.No ratings yet
- Cats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisFrom EverandCats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisNo ratings yet
- K-4 PR - Soil Transmitted HelminthiasisDocument45 pagesK-4 PR - Soil Transmitted HelminthiasissantayohanaNo ratings yet
- SCORE Studies On The Impact of Drug Treatment On Morbidity Due To Schistosoma Mansoni and Schistosoma Haematobium InfectionDocument6 pagesSCORE Studies On The Impact of Drug Treatment On Morbidity Due To Schistosoma Mansoni and Schistosoma Haematobium InfectionAneela HereNo ratings yet
- Typhoid Fever PDFDocument14 pagesTyphoid Fever PDFginstarNo ratings yet
- Ascaris y AscaridiasisDocument6 pagesAscaris y Ascaridiasisemmanueltobon0621No ratings yet
- Hemolytic-Uremic SyndromeDocument16 pagesHemolytic-Uremic Syndromeabdel HigazyNo ratings yet
- Clinical Considerations On Canine Visceral Leishmaniasis in Greece: A Retrospective Study of 158 Cases (1989-1996)Document8 pagesClinical Considerations On Canine Visceral Leishmaniasis in Greece: A Retrospective Study of 158 Cases (1989-1996)5-Mint PUBGNo ratings yet
- Mbio 01490-14Document9 pagesMbio 01490-14Rin ChanNo ratings yet
- Schistosoma Mansoni Treatment Reduces HIV Entry Into Cervical CD4 + T Cells and Induces IFN-I PathwaysDocument12 pagesSchistosoma Mansoni Treatment Reduces HIV Entry Into Cervical CD4 + T Cells and Induces IFN-I PathwaysMariaIzabelLopesNo ratings yet
- Heligmosmoides ArthritisDocument8 pagesHeligmosmoides ArthritisCagar Irwin TaufanNo ratings yet
- Final Project VinceDocument21 pagesFinal Project Vincekiss4444f4No ratings yet
- Metronidazole Resistance Caco 2 AdhesionDocument18 pagesMetronidazole Resistance Caco 2 AdhesionDyego CarléttiNo ratings yet
- Morton 2005Document23 pagesMorton 2005Bernardo AlmeidaNo ratings yet
- NIH Public Access: Author ManuscriptDocument20 pagesNIH Public Access: Author ManuscriptAdriana OrtizNo ratings yet
- Effects of Iron Chelating Agent On Schistosoma Mans - 2019 - Biomedicine - PharmDocument11 pagesEffects of Iron Chelating Agent On Schistosoma Mans - 2019 - Biomedicine - PharmPryscila Zanetti OmsNo ratings yet
- Acute Osteomyelitis in ChildrenDocument12 pagesAcute Osteomyelitis in ChildrenSari MuranasNo ratings yet
- Negrini2021 Article SjögrenSSyndromeASystemicAutoiDocument17 pagesNegrini2021 Article SjögrenSSyndromeASystemicAutoiSahanaNo ratings yet
- Endemic Mycoses: Blastomycosis, Histoplasmosis, and SporotrichosisDocument18 pagesEndemic Mycoses: Blastomycosis, Histoplasmosis, and SporotrichosisaugustosavioNo ratings yet
- Diseases 07 00028Document14 pagesDiseases 07 00028shaznay delacruzNo ratings yet
- Pneumocystis Jiroveci: Jay A. Fishman, MDDocument17 pagesPneumocystis Jiroveci: Jay A. Fishman, MDIsmael Armando Collantes DiazNo ratings yet
- Mullangi 2012Document10 pagesMullangi 2012Mariana AmbossNo ratings yet
- Harrus 1999Document5 pagesHarrus 1999Paula Andrea Castro MartinezNo ratings yet
- 1 s2.0 S0165587618305354 MainDocument6 pages1 s2.0 S0165587618305354 MainChrista24796No ratings yet
- Intestinal HelminthsDocument4 pagesIntestinal HelminthssivaNo ratings yet
- Knowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaDocument6 pagesKnowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaAbdirahman YazinNo ratings yet
- Adaptation To The Host and Persistence: Role of Loss of Capsular Polysaccharide ExpressionDocument10 pagesAdaptation To The Host and Persistence: Role of Loss of Capsular Polysaccharide ExpressionIslam ZhyanmaNo ratings yet
- Prevention and Control of Amoebic DysenteryDocument16 pagesPrevention and Control of Amoebic DysenteryPakSci MissionNo ratings yet
- PDM Os 9Document15 pagesPDM Os 9Erdinawira RizkianiNo ratings yet
- Aspergillosis FinalDocument39 pagesAspergillosis FinalBhupendraNo ratings yet
- Inflammation: Cardinal Signs of InflammationDocument9 pagesInflammation: Cardinal Signs of InflammationGeoffreyNo ratings yet
- Acute Colitis DiagnosisDocument23 pagesAcute Colitis Diagnosismarielahuerta123No ratings yet
- Palatal Perforation: Case Report and Literature Review: 10.5005/jp-Journals-10039-1170Document4 pagesPalatal Perforation: Case Report and Literature Review: 10.5005/jp-Journals-10039-1170Maqbul AlamNo ratings yet
- Candidates Mcqs Pathology Week 11: 1 JF A BDocument3 pagesCandidates Mcqs Pathology Week 11: 1 JF A Bمجتبى علي0% (1)
- Patogenesis TBDocument46 pagesPatogenesis TBJaya Semara PutraNo ratings yet
- Non-Neoplastic Disorders: Lymphoid SystemDocument36 pagesNon-Neoplastic Disorders: Lymphoid SystemMutiana Muspita JeliNo ratings yet
- Pathology Week 2 p1-18Document18 pagesPathology Week 2 p1-18zeroun24100% (1)
- Pathogenesis and Morphorlogy of Tuberculosis M. O. UdohDocument6 pagesPathogenesis and Morphorlogy of Tuberculosis M. O. UdohBelleNo ratings yet
- History: Tuberculosis (TB)Document24 pagesHistory: Tuberculosis (TB)Rashed ShatnawiNo ratings yet
- Sheffield Diagnostic Histopathology Course Soft Tissue TumoursDocument42 pagesSheffield Diagnostic Histopathology Course Soft Tissue TumoursSimionescu FlorentinaNo ratings yet
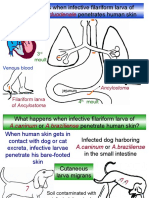
- Ancylostoma Duodenale: What Happens When Infective Filariform Larva of Penetrates Human Skin ?Document16 pagesAncylostoma Duodenale: What Happens When Infective Filariform Larva of Penetrates Human Skin ?Ayop KhNo ratings yet
- Patho Lec Block 4Document35 pagesPatho Lec Block 4Dale OcampoNo ratings yet
- Metal and The SkinDocument441 pagesMetal and The SkinAGUSTONI pUJIATO100% (2)
- Self Assessment CasesDocument512 pagesSelf Assessment CasesmixandgoNo ratings yet
- Test - 9 Chronic PeriodontitisDocument5 pagesTest - 9 Chronic PeriodontitisIsak ShatikaNo ratings yet
- SESLHDPR 400 - Wound Management of Hypergranulation Tissue - 0Document7 pagesSESLHDPR 400 - Wound Management of Hypergranulation Tissue - 0MohammadNo ratings yet
- Stomach - GastritisDocument22 pagesStomach - GastritisaimanNo ratings yet
- Unit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsDocument85 pagesUnit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsCherry RahimaNo ratings yet
- Panniculitis 2021Document45 pagesPanniculitis 2021karimahihdaNo ratings yet
- Medically Important Bacteria Memorisation SheetDocument7 pagesMedically Important Bacteria Memorisation SheetOlivia LimNo ratings yet
- Fayara Aretha Kunaefi Pathophysiology QBD1 FG2Document25 pagesFayara Aretha Kunaefi Pathophysiology QBD1 FG2fayaNo ratings yet
- Granulomatous Colitis of Boxer DogsDocument13 pagesGranulomatous Colitis of Boxer DogsEduardo SantamaríaNo ratings yet
- Ciri Khas Histo PADocument28 pagesCiri Khas Histo PAFitria SartikaNo ratings yet
- Clinicopathological Spectrum of Benign Oral Cavity LesionsDocument8 pagesClinicopathological Spectrum of Benign Oral Cavity LesionsIJAR JOURNALNo ratings yet
- A Foreign Body Reaction To Surgicel R in A Lymph NDocument2 pagesA Foreign Body Reaction To Surgicel R in A Lymph NIndustrijsko MasinskiNo ratings yet

























































































Download as pdf or txt
You might also like
- High Yield NBME Images AnnotatedDocument84 pagesHigh Yield NBME Images AnnotatedNareman Abd El HadeNo ratings yet
- Diagnosis of Small Lung Biopsy An Integrated Approach by Mostafa M. FraigDocument111 pagesDiagnosis of Small Lung Biopsy An Integrated Approach by Mostafa M. FraigFlaviaNo ratings yet
- (Medical Masterclass) Coll.-Infectious Diseases and DermatologyDocument333 pages(Medical Masterclass) Coll.-Infectious Diseases and DermatologyVijay Mg100% (2)
- Microscopic Slides Pathology-2Document135 pagesMicroscopic Slides Pathology-2Justin JannatiNo ratings yet
- Immune Response To SchistosomaDocument11 pagesImmune Response To SchistosomaRobMarvinNo ratings yet
- Blastocytosis Hominis - The Summary From A To ZDocument11 pagesBlastocytosis Hominis - The Summary From A To ZSalsabila TasyaNo ratings yet
- Schistosoma "Eggs-Iting" The Host: Granuloma Formation and Egg ExcretionDocument16 pagesSchistosoma "Eggs-Iting" The Host: Granuloma Formation and Egg ExcretionRoberto Evangelista CostaNo ratings yet
- Schistosomiasis and Liver Fibrosis: Review ArticleDocument8 pagesSchistosomiasis and Liver Fibrosis: Review ArticlevigneshNo ratings yet
- Jordan University Faculty of Dentistry DR Mohammad Al-Tamimi, MD, PHDDocument20 pagesJordan University Faculty of Dentistry DR Mohammad Al-Tamimi, MD, PHDDaniel AtiehNo ratings yet
- PatogenesisDocument28 pagesPatogenesisOka WidyaningsihNo ratings yet
- Fimmu 04 00507 PDFDocument20 pagesFimmu 04 00507 PDFSilma FarrahaNo ratings yet
- BMJ d2651 PDFDocument11 pagesBMJ d2651 PDFKarin KurniatiNo ratings yet
- Key Infections in The PlacentaDocument14 pagesKey Infections in The PlacentaLauraGonzalezNo ratings yet
- Schistosomiasis: ArticleDocument20 pagesSchistosomiasis: ArticleVincent ReyesNo ratings yet
- Salmonella, The Host and Disease: A Brief ReviewDocument8 pagesSalmonella, The Host and Disease: A Brief ReviewPedro Albán MNo ratings yet
- 2015 Kline Bowdish Infections in An Aging PopulationDocument5 pages2015 Kline Bowdish Infections in An Aging PopulationLydiaphNo ratings yet
- Jiu 189Document9 pagesJiu 189barreirosanchez984No ratings yet
- Petton 2021 Frontiers Pacific MANUSCRIT2Document26 pagesPetton 2021 Frontiers Pacific MANUSCRIT2AmadorRevillaNo ratings yet
- Gram-Positive Cocci Staphylococcus CLINICAL SIGNIFICANCEDocument8 pagesGram-Positive Cocci Staphylococcus CLINICAL SIGNIFICANCEsobia faryadaliNo ratings yet
- Advances in New Target Molecules Against - Peng ZhuDocument22 pagesAdvances in New Target Molecules Against - Peng Zhuana.pinho.souzaNo ratings yet
- Human SchistosomiasisDocument18 pagesHuman SchistosomiasisbassbngNo ratings yet
- Othman 2014Document44 pagesOthman 2014Jessica QuevedoNo ratings yet
- 6 Helminth2 - PPTDocument8 pages6 Helminth2 - PPTRaihan Zulfiqar HalimNo ratings yet
- Arch Dis Child 1998 Ladhani 85 8Document5 pagesArch Dis Child 1998 Ladhani 85 8Dian Ayu Permata SandiNo ratings yet
- Blastocystis How Do Specific Diets and Human Gut MicrobiotaDocument10 pagesBlastocystis How Do Specific Diets and Human Gut MicrobiotaVanessa OcegueraNo ratings yet
- Empyema 2Document9 pagesEmpyema 2phobicmdNo ratings yet
- Lyon 2005Document21 pagesLyon 2005Renata StefaniNo ratings yet
- At The Turn of The 20th CenturyDocument6 pagesAt The Turn of The 20th CenturyStefanosSokratousNo ratings yet
- Location and Pathogenic Potenctial of Blastocystis in The Porcine IntestineDocument8 pagesLocation and Pathogenic Potenctial of Blastocystis in The Porcine Intestinesebasrd07No ratings yet
- Major Serological Screening Methodologies To Anti - T. Gondii Antibodies Detection in Pregnancy: Mini Literature ViewDocument4 pagesMajor Serological Screening Methodologies To Anti - T. Gondii Antibodies Detection in Pregnancy: Mini Literature Viewpublishing groupNo ratings yet
- Pancreatic Tuberculosis Review ArticleDocument24 pagesPancreatic Tuberculosis Review ArticlesamudraandiNo ratings yet
- Isospora SuisDocument35 pagesIsospora SuisAfif DihanNo ratings yet
- Interactions Between Parasites and Human MicrobiotDocument6 pagesInteractions Between Parasites and Human MicrobiotGregorio De Las CasasNo ratings yet
- Toxo PregnancyDocument15 pagesToxo PregnancyYossy CatarinaNo ratings yet
- First Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDocument11 pagesFirst Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDavid A. Osorio G.No ratings yet
- Cats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisFrom EverandCats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisNo ratings yet
- K-4 PR - Soil Transmitted HelminthiasisDocument45 pagesK-4 PR - Soil Transmitted HelminthiasissantayohanaNo ratings yet
- SCORE Studies On The Impact of Drug Treatment On Morbidity Due To Schistosoma Mansoni and Schistosoma Haematobium InfectionDocument6 pagesSCORE Studies On The Impact of Drug Treatment On Morbidity Due To Schistosoma Mansoni and Schistosoma Haematobium InfectionAneela HereNo ratings yet
- Typhoid Fever PDFDocument14 pagesTyphoid Fever PDFginstarNo ratings yet
- Ascaris y AscaridiasisDocument6 pagesAscaris y Ascaridiasisemmanueltobon0621No ratings yet
- Hemolytic-Uremic SyndromeDocument16 pagesHemolytic-Uremic Syndromeabdel HigazyNo ratings yet
- Clinical Considerations On Canine Visceral Leishmaniasis in Greece: A Retrospective Study of 158 Cases (1989-1996)Document8 pagesClinical Considerations On Canine Visceral Leishmaniasis in Greece: A Retrospective Study of 158 Cases (1989-1996)5-Mint PUBGNo ratings yet
- Mbio 01490-14Document9 pagesMbio 01490-14Rin ChanNo ratings yet
- Schistosoma Mansoni Treatment Reduces HIV Entry Into Cervical CD4 + T Cells and Induces IFN-I PathwaysDocument12 pagesSchistosoma Mansoni Treatment Reduces HIV Entry Into Cervical CD4 + T Cells and Induces IFN-I PathwaysMariaIzabelLopesNo ratings yet
- Heligmosmoides ArthritisDocument8 pagesHeligmosmoides ArthritisCagar Irwin TaufanNo ratings yet
- Final Project VinceDocument21 pagesFinal Project Vincekiss4444f4No ratings yet
- Metronidazole Resistance Caco 2 AdhesionDocument18 pagesMetronidazole Resistance Caco 2 AdhesionDyego CarléttiNo ratings yet
- Morton 2005Document23 pagesMorton 2005Bernardo AlmeidaNo ratings yet
- NIH Public Access: Author ManuscriptDocument20 pagesNIH Public Access: Author ManuscriptAdriana OrtizNo ratings yet
- Effects of Iron Chelating Agent On Schistosoma Mans - 2019 - Biomedicine - PharmDocument11 pagesEffects of Iron Chelating Agent On Schistosoma Mans - 2019 - Biomedicine - PharmPryscila Zanetti OmsNo ratings yet
- Acute Osteomyelitis in ChildrenDocument12 pagesAcute Osteomyelitis in ChildrenSari MuranasNo ratings yet
- Negrini2021 Article SjögrenSSyndromeASystemicAutoiDocument17 pagesNegrini2021 Article SjögrenSSyndromeASystemicAutoiSahanaNo ratings yet
- Endemic Mycoses: Blastomycosis, Histoplasmosis, and SporotrichosisDocument18 pagesEndemic Mycoses: Blastomycosis, Histoplasmosis, and SporotrichosisaugustosavioNo ratings yet
- Diseases 07 00028Document14 pagesDiseases 07 00028shaznay delacruzNo ratings yet
- Pneumocystis Jiroveci: Jay A. Fishman, MDDocument17 pagesPneumocystis Jiroveci: Jay A. Fishman, MDIsmael Armando Collantes DiazNo ratings yet
- Mullangi 2012Document10 pagesMullangi 2012Mariana AmbossNo ratings yet
- Harrus 1999Document5 pagesHarrus 1999Paula Andrea Castro MartinezNo ratings yet
- 1 s2.0 S0165587618305354 MainDocument6 pages1 s2.0 S0165587618305354 MainChrista24796No ratings yet
- Intestinal HelminthsDocument4 pagesIntestinal HelminthssivaNo ratings yet
- Knowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaDocument6 pagesKnowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaAbdirahman YazinNo ratings yet
- Adaptation To The Host and Persistence: Role of Loss of Capsular Polysaccharide ExpressionDocument10 pagesAdaptation To The Host and Persistence: Role of Loss of Capsular Polysaccharide ExpressionIslam ZhyanmaNo ratings yet
- Prevention and Control of Amoebic DysenteryDocument16 pagesPrevention and Control of Amoebic DysenteryPakSci MissionNo ratings yet
- PDM Os 9Document15 pagesPDM Os 9Erdinawira RizkianiNo ratings yet
- Aspergillosis FinalDocument39 pagesAspergillosis FinalBhupendraNo ratings yet
- Inflammation: Cardinal Signs of InflammationDocument9 pagesInflammation: Cardinal Signs of InflammationGeoffreyNo ratings yet
- Acute Colitis DiagnosisDocument23 pagesAcute Colitis Diagnosismarielahuerta123No ratings yet
- Palatal Perforation: Case Report and Literature Review: 10.5005/jp-Journals-10039-1170Document4 pagesPalatal Perforation: Case Report and Literature Review: 10.5005/jp-Journals-10039-1170Maqbul AlamNo ratings yet
- Candidates Mcqs Pathology Week 11: 1 JF A BDocument3 pagesCandidates Mcqs Pathology Week 11: 1 JF A Bمجتبى علي0% (1)
- Patogenesis TBDocument46 pagesPatogenesis TBJaya Semara PutraNo ratings yet
- Non-Neoplastic Disorders: Lymphoid SystemDocument36 pagesNon-Neoplastic Disorders: Lymphoid SystemMutiana Muspita JeliNo ratings yet
- Pathology Week 2 p1-18Document18 pagesPathology Week 2 p1-18zeroun24100% (1)
- Pathogenesis and Morphorlogy of Tuberculosis M. O. UdohDocument6 pagesPathogenesis and Morphorlogy of Tuberculosis M. O. UdohBelleNo ratings yet
- History: Tuberculosis (TB)Document24 pagesHistory: Tuberculosis (TB)Rashed ShatnawiNo ratings yet
- Sheffield Diagnostic Histopathology Course Soft Tissue TumoursDocument42 pagesSheffield Diagnostic Histopathology Course Soft Tissue TumoursSimionescu FlorentinaNo ratings yet
- Ancylostoma Duodenale: What Happens When Infective Filariform Larva of Penetrates Human Skin ?Document16 pagesAncylostoma Duodenale: What Happens When Infective Filariform Larva of Penetrates Human Skin ?Ayop KhNo ratings yet
- Patho Lec Block 4Document35 pagesPatho Lec Block 4Dale OcampoNo ratings yet
- Metal and The SkinDocument441 pagesMetal and The SkinAGUSTONI pUJIATO100% (2)
- Self Assessment CasesDocument512 pagesSelf Assessment CasesmixandgoNo ratings yet
- Test - 9 Chronic PeriodontitisDocument5 pagesTest - 9 Chronic PeriodontitisIsak ShatikaNo ratings yet
- SESLHDPR 400 - Wound Management of Hypergranulation Tissue - 0Document7 pagesSESLHDPR 400 - Wound Management of Hypergranulation Tissue - 0MohammadNo ratings yet
- Stomach - GastritisDocument22 pagesStomach - GastritisaimanNo ratings yet
- Unit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsDocument85 pagesUnit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsCherry RahimaNo ratings yet
- Panniculitis 2021Document45 pagesPanniculitis 2021karimahihdaNo ratings yet
- Medically Important Bacteria Memorisation SheetDocument7 pagesMedically Important Bacteria Memorisation SheetOlivia LimNo ratings yet
- Fayara Aretha Kunaefi Pathophysiology QBD1 FG2Document25 pagesFayara Aretha Kunaefi Pathophysiology QBD1 FG2fayaNo ratings yet
- Granulomatous Colitis of Boxer DogsDocument13 pagesGranulomatous Colitis of Boxer DogsEduardo SantamaríaNo ratings yet
- Ciri Khas Histo PADocument28 pagesCiri Khas Histo PAFitria SartikaNo ratings yet
- Clinicopathological Spectrum of Benign Oral Cavity LesionsDocument8 pagesClinicopathological Spectrum of Benign Oral Cavity LesionsIJAR JOURNALNo ratings yet
- A Foreign Body Reaction To Surgicel R in A Lymph NDocument2 pagesA Foreign Body Reaction To Surgicel R in A Lymph NIndustrijsko MasinskiNo ratings yet