Download as pdf or txt
You might also like
- The Art of Accompanying (Classic Reprint) - LINDO, Algernon H - PDFDocument121 pagesThe Art of Accompanying (Classic Reprint) - LINDO, Algernon H - PDFDimitrios Sykias100% (4)
- Case Incident OB MBADocument4 pagesCase Incident OB MBASahar Al-Jobury57% (7)
- The Brain's Default Network and Its Adaptive Role in InternalDocument29 pagesThe Brain's Default Network and Its Adaptive Role in Internaldarketa100% (1)
- Unveiling Complexity: Non-Linear and Fractal Analysis in Neuroscience and Cognitive PsychologyDocument2 pagesUnveiling Complexity: Non-Linear and Fractal Analysis in Neuroscience and Cognitive PsychologyNicky KellyNo ratings yet
- Topological Exploration of Artificial Neuronal Network DynamicsDocument20 pagesTopological Exploration of Artificial Neuronal Network DynamicsYishuai ZhangNo ratings yet
- Neuroscience of Psychoanalytic Object Relations Theory MineDocument59 pagesNeuroscience of Psychoanalytic Object Relations Theory MineTatjana Jočić VelimirovićNo ratings yet
- The Brain's Default Network: Anatomy, Function, and Relevance To DiseaseDocument38 pagesThe Brain's Default Network: Anatomy, Function, and Relevance To DiseaseMarkoff ChaneyNo ratings yet
- EEG Hyperscanning in Motor Rehabilitation, A Position PaperDocument15 pagesEEG Hyperscanning in Motor Rehabilitation, A Position PaperWarnithaNo ratings yet
- Functional Organization of Intrinsic Connectivity Networks in Chinese-Chess ExpertsDocument11 pagesFunctional Organization of Intrinsic Connectivity Networks in Chinese-Chess ExpertsAlambre PúaNo ratings yet
- NIH Public Access: Future Trends in Neuroimaging: Neural Processes As Expressed Within Real-Life ContextsDocument14 pagesNIH Public Access: Future Trends in Neuroimaging: Neural Processes As Expressed Within Real-Life ContextslitrevnmNo ratings yet
- Exploring The Collective Brain ActivityDocument30 pagesExploring The Collective Brain Activitynikolas decoNo ratings yet
- Do We Need To Study The Brain To Understand The MindDocument3 pagesDo We Need To Study The Brain To Understand The MindMARCO ANTONIO OLIVARES HURTADONo ratings yet
- Keshmiri 2022Document24 pagesKeshmiri 2022Berit SingerNo ratings yet
- Encoding and Decoding Models in Cognitive ElectrophysiologyDocument24 pagesEncoding and Decoding Models in Cognitive ElectrophysiologyOak TreeNo ratings yet
- Coherence: The Measurement and Application of Brain ConnectivityDocument12 pagesCoherence: The Measurement and Application of Brain ConnectivitySekretariat Neurologi UNPADNo ratings yet
- Review: Conscious Processing and The Global Neuronal Workspace HypothesisDocument23 pagesReview: Conscious Processing and The Global Neuronal Workspace HypothesisSebastian Gonzalez MazoNo ratings yet
- Burgess 08 SpatialDocument21 pagesBurgess 08 Spatialsarabiba1971No ratings yet
- Cognition AlgorithmDocument19 pagesCognition AlgorithmThiago RibeiroNo ratings yet
- Quantum Physics in Neuroscience and PsychologyDocument19 pagesQuantum Physics in Neuroscience and PsychologySabrina MorelliNo ratings yet
- 2004 Towards Studies of The Social BrainDocument5 pages2004 Towards Studies of The Social BrainAl WilsonNo ratings yet
- Understanding Neural Exibility From A Multifaceted de NitionDocument16 pagesUnderstanding Neural Exibility From A Multifaceted de Nition童帅No ratings yet
- An Integrated Index Engrams Place Cells and Hippocampal Memory Travis D Goode Kazumasa Z Tanaka Amar Sahay Thomas J Mchugh Full ChapterDocument35 pagesAn Integrated Index Engrams Place Cells and Hippocampal Memory Travis D Goode Kazumasa Z Tanaka Amar Sahay Thomas J Mchugh Full Chapterandrea.davis765100% (6)
- (Bettcher Et Al., 2016) PDFDocument35 pages(Bettcher Et Al., 2016) PDFFARHAT HAJERNo ratings yet
- Articulo InglesDocument20 pagesArticulo InglesPedro Coronado ArrascueNo ratings yet
- MMosco Gilboa - Handbook - Final - Dec23Document125 pagesMMosco Gilboa - Handbook - Final - Dec23mirelle.acNo ratings yet
- Intelligence and Tuning inDocument17 pagesIntelligence and Tuning inBaltazar alvesNo ratings yet
- HHS Public Access: Neural Architecture Underlying Classification of Face Perception ParadigmsDocument26 pagesHHS Public Access: Neural Architecture Underlying Classification of Face Perception Paradigmscecilia martinezNo ratings yet
- Quiroga-Lombardetal. J.Neurophysiol. 2013Document12 pagesQuiroga-Lombardetal. J.Neurophysiol. 2013Vinicius de SousaNo ratings yet
- 1 s2.0 S0010945221002409 MainDocument16 pages1 s2.0 S0010945221002409 Mainelbueno21No ratings yet
- Nonlinear Dynamics of The Brain: Emotion and Cognition: Home Search Collections Journals About Contact Us My IopscienceDocument17 pagesNonlinear Dynamics of The Brain: Emotion and Cognition: Home Search Collections Journals About Contact Us My Iopsciencebrinkman94No ratings yet
- NIH Public Access: Multivariate Pattern Analysis of fMRI: The Early BeginningsDocument7 pagesNIH Public Access: Multivariate Pattern Analysis of fMRI: The Early BeginningsfiestamixNo ratings yet
- Brain Mechanisms of Concept LearningDocument8 pagesBrain Mechanisms of Concept LearningCarlos AlmeidaNo ratings yet
- 16848-Article Text-68192-2-10-20170313Document11 pages16848-Article Text-68192-2-10-20170313Yosi KusukaNo ratings yet
- 1 Functional Neuroimaging of Executive Functions Todd S. Braver and Hannes Ruge Department ... (PDFDrive)Document93 pages1 Functional Neuroimaging of Executive Functions Todd S. Braver and Hannes Ruge Department ... (PDFDrive)Amornchai WongworrakunNo ratings yet
- Dynamical Exploration of The Repertoire of Brain Networks at Rest Is Modulated by PsilocybinDocument16 pagesDynamical Exploration of The Repertoire of Brain Networks at Rest Is Modulated by PsilocybinDayon BochniaNo ratings yet
- Putational Neuroscience - HandbookDocument26 pagesPutational Neuroscience - Handbookdetki007No ratings yet
- Neuroscience-Inspired Artificial IntelligenceDocument15 pagesNeuroscience-Inspired Artificial IntelligenceGOGUNo ratings yet
- Van Der Houvel, 2011 Rich Club NetworkDocument12 pagesVan Der Houvel, 2011 Rich Club NetworkRicardo Jose De LeonNo ratings yet
- Aphasia RehabDocument72 pagesAphasia RehabkicsirekaNo ratings yet
- NATUREDocument12 pagesNATUREGuadalupe BerbigierNo ratings yet
- Quantum Physics in Neuroscience and Psychology - A New Theory With Respect To Mind - Brain Interacti PDFDocument72 pagesQuantum Physics in Neuroscience and Psychology - A New Theory With Respect To Mind - Brain Interacti PDFAlina Petrut100% (1)
- Quantum Physics in Neuroscience and PsychologyDocument20 pagesQuantum Physics in Neuroscience and PsychologyPeter SutorNo ratings yet
- Free Neuro ArticleDocument15 pagesFree Neuro ArticleShafee MohammedNo ratings yet
- Coordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorFrom EverandCoordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorJose Luis Perez VelazquezNo ratings yet
- Editorial: J.Neurol Sci (Turk)Document4 pagesEditorial: J.Neurol Sci (Turk)Nezih OktarNo ratings yet
- Simplicial and Topological Descriptions of Human Brain DynamicsDocument20 pagesSimplicial and Topological Descriptions of Human Brain DynamicsJosue VasquezNo ratings yet
- Plasticité Nrpsy CogDocument14 pagesPlasticité Nrpsy CogMalak GabsiNo ratings yet
- Human Brain Mapping - 2005 - Patel - A Bayesian Approach To Determining Connectivity of The Human BrainDocument10 pagesHuman Brain Mapping - 2005 - Patel - A Bayesian Approach To Determining Connectivity of The Human BrainKerem YücesanNo ratings yet
- Conect FX AmigdalaDocument47 pagesConect FX AmigdalamarielaNo ratings yet
- MVPA Personality - PreprintDocument14 pagesMVPA Personality - PreprintGeo MeNo ratings yet
- tmp1DBF TMPDocument3 pagestmp1DBF TMPFrontiersNo ratings yet
- Brain Activity Hypnosis Study PDFDocument11 pagesBrain Activity Hypnosis Study PDFDavid OcNo ratings yet
- The Maturing Architecture of The Brain's Default NetworkDocument5 pagesThe Maturing Architecture of The Brain's Default Networkমেঘে ঢাকা তারাNo ratings yet
- Soc Cogn Affect Neurosci 2016 Greven 641 51Document11 pagesSoc Cogn Affect Neurosci 2016 Greven 641 51tefsNo ratings yet
- Cirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesCirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDanaNo ratings yet
- Brain Functioning and Regeneration: Kinematics of the Brain Activities Volume IvFrom EverandBrain Functioning and Regeneration: Kinematics of the Brain Activities Volume IvNo ratings yet
- Kinematics of the Brain Activities Vol. V: Plasticity, Elasticity and Resonating of the Neural NetworksFrom EverandKinematics of the Brain Activities Vol. V: Plasticity, Elasticity and Resonating of the Neural NetworksNo ratings yet
- Semblance Hypothesis of Memory: 3Rd EditionFrom EverandSemblance Hypothesis of Memory: 3Rd EditionRating: 5 out of 5 stars5/5 (1)
- Arrington - Logan 2005Document21 pagesArrington - Logan 2005Francisco Ahumada MéndezNo ratings yet
- 1 - Leeuw2023Document8 pages1 - Leeuw2023Francisco Ahumada MéndezNo ratings yet
- Arrington2008Document7 pagesArrington2008Francisco Ahumada MéndezNo ratings yet
- Arrington - Logan 2004Document7 pagesArrington - Logan 2004Francisco Ahumada MéndezNo ratings yet
- Liefooghe2009Document8 pagesLiefooghe2009Francisco Ahumada MéndezNo ratings yet
- Yu Chin Chiu2020Document8 pagesYu Chin Chiu2020Francisco Ahumada MéndezNo ratings yet
- Mayr2006Document7 pagesMayr2006Francisco Ahumada MéndezNo ratings yet
- Mood Induction Effects On Motor Sequence Learning and Stop Signal Reaction TimeDocument16 pagesMood Induction Effects On Motor Sequence Learning and Stop Signal Reaction TimeFrancisco Ahumada MéndezNo ratings yet
- Arrington2009Document6 pagesArrington2009Francisco Ahumada MéndezNo ratings yet
- PRISMA PAGE 2020 ExplanationDocument36 pagesPRISMA PAGE 2020 ExplanationFrancisco Ahumada MéndezNo ratings yet
- Seeking Optimal Confusion: A Review On Epistemic Emotion Management in Interactive Digital Learning EnvironmentsDocument12 pagesSeeking Optimal Confusion: A Review On Epistemic Emotion Management in Interactive Digital Learning EnvironmentsFrancisco Ahumada MéndezNo ratings yet
- Barrett 2010Document30 pagesBarrett 2010Francisco Ahumada MéndezNo ratings yet
- Cognition: A A B ADocument7 pagesCognition: A A B AFrancisco Ahumada MéndezNo ratings yet
- From Conditioning To Emotion: Translating Animal Models of Learning To Human PsychopathologyDocument14 pagesFrom Conditioning To Emotion: Translating Animal Models of Learning To Human PsychopathologyFrancisco Ahumada MéndezNo ratings yet
- Computers in Human Behavior: Quan Chen, Zheng YanDocument11 pagesComputers in Human Behavior: Quan Chen, Zheng YanFrancisco Ahumada MéndezNo ratings yet
- LinguaPix Database 2021Document14 pagesLinguaPix Database 2021Francisco Ahumada MéndezNo ratings yet
- Zhang2016Document10 pagesZhang2016Francisco Ahumada MéndezNo ratings yet
- 31 - Vanlessen2013Document20 pages31 - Vanlessen2013Francisco Ahumada MéndezNo ratings yet
- Evidence For A Multicomponent Hierarchical Representation of Dual TasksDocument14 pagesEvidence For A Multicomponent Hierarchical Representation of Dual TasksFrancisco Ahumada MéndezNo ratings yet
- 1 - Wacker2017Document16 pages1 - Wacker2017Francisco Ahumada MéndezNo ratings yet
- The Urban Security Image Database (USID)Document21 pagesThe Urban Security Image Database (USID)Francisco Ahumada MéndezNo ratings yet
- 3 - Isiea2021Document13 pages3 - Isiea2021Francisco Ahumada MéndezNo ratings yet
- Revisiting Positive Affect and Reward Influences On Cognitive ControlDocument7 pagesRevisiting Positive Affect and Reward Influences On Cognitive ControlFrancisco Ahumada MéndezNo ratings yet
- Suppression (But Not Reappraisal) Impairs Subsequent Error Detection: An ERP Study of Emotion Regulation's Resource-Depleting EffectDocument9 pagesSuppression (But Not Reappraisal) Impairs Subsequent Error Detection: An ERP Study of Emotion Regulation's Resource-Depleting EffectFrancisco Ahumada MéndezNo ratings yet
- Arousal (But Not Valence) Amplifies The Impact of Salience: Cognition and EmotionDocument8 pagesArousal (But Not Valence) Amplifies The Impact of Salience: Cognition and EmotionFrancisco Ahumada MéndezNo ratings yet
- Arousal-Biased Competition in Perception and Memory: Mara Mather and Matthew R. SutherlandDocument20 pagesArousal-Biased Competition in Perception and Memory: Mara Mather and Matthew R. SutherlandFrancisco Ahumada MéndezNo ratings yet
- 7 - Straub2019valenceDocument16 pages7 - Straub2019valenceFrancisco Ahumada MéndezNo ratings yet
- Joseph P. Forgas - Handbook of Affect and Social Cognition-Taylor & Francis (2001)Document612 pagesJoseph P. Forgas - Handbook of Affect and Social Cognition-Taylor & Francis (2001)Francisco Ahumada MéndezNo ratings yet
- Neuropsychologia: Thomas Goschke, Annette BolteDocument21 pagesNeuropsychologia: Thomas Goschke, Annette BolteFrancisco Ahumada MéndezNo ratings yet
- Dignath2020phasic TonicDocument24 pagesDignath2020phasic TonicFrancisco Ahumada MéndezNo ratings yet
- FRC Essays - Comply or ExplainDocument64 pagesFRC Essays - Comply or ExplainRahat TabassumNo ratings yet
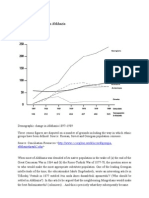
- Demographic Change in AbkhaziaDocument3 pagesDemographic Change in AbkhaziacircassianworldNo ratings yet
- Jekyll and Hyde Interesting ContextDocument6 pagesJekyll and Hyde Interesting Contextreece.dillon01No ratings yet
- Drug Study: Tabije, Arvie Jayselle P. Marbella, IvyDocument15 pagesDrug Study: Tabije, Arvie Jayselle P. Marbella, IvyJaysellePuguonTabijeNo ratings yet
- Performing Short - 1Document4 pagesPerforming Short - 1mshahidshaukatNo ratings yet
- NCP PryllDocument6 pagesNCP PryllpjcolitaNo ratings yet
- Resume Brandon Stanyer 2022Document2 pagesResume Brandon Stanyer 2022api-602194418No ratings yet
- C & D Series Backhoe Loader Diagnostics: Service TrainingDocument25 pagesC & D Series Backhoe Loader Diagnostics: Service TrainingFaustoNo ratings yet
- Screen Time Final EssayDocument5 pagesScreen Time Final EssayAlisha PowellNo ratings yet
- Continuous Beam Staad ReportDocument14 pagesContinuous Beam Staad ReportMuhammad Shukri Bin SaharinNo ratings yet
- Barishal Project ProposalDocument7 pagesBarishal Project Proposaltipu1sultan_1No ratings yet
- Colpitts OscillatorDocument4 pagesColpitts OscillatorAnimesh Bhanja100% (1)
- Literature Concepts Terms: Familiarize YourselfDocument25 pagesLiterature Concepts Terms: Familiarize YourselfPheobie Sheela V. CuartocruzNo ratings yet
- 2017 - Orientalism and The Qur Ān A ProlegomenonDocument8 pages2017 - Orientalism and The Qur Ān A ProlegomenonMohammad -No ratings yet
- EMBA Applied Value Investing (Ajdler) SP2016Document3 pagesEMBA Applied Value Investing (Ajdler) SP2016darwin12No ratings yet
- Advertising of A Beauty Parlour BusinessDocument4 pagesAdvertising of A Beauty Parlour Businessjerrypassah09No ratings yet
- Uea Coursework Word CountDocument4 pagesUea Coursework Word Counttgpazszid100% (2)
- Course Outline - DEA 2313Document11 pagesCourse Outline - DEA 2313Roger JohnNo ratings yet
- Hem Lab Manual DiffDocument9 pagesHem Lab Manual DiffFatima Mae LusanNo ratings yet
- PMP Test Paper 101 Questions For 2hours PDFDocument21 pagesPMP Test Paper 101 Questions For 2hours PDFAnonymous 6pfaIAh8100% (1)
- Planning Your Trades: Risk Management RiskDocument2 pagesPlanning Your Trades: Risk Management RiskdoremonNo ratings yet
- Strategic Mindset - Action GuideDocument19 pagesStrategic Mindset - Action GuideTinki DewanganNo ratings yet
- Few Good Men AssignmentDocument3 pagesFew Good Men AssignmentBellaNo ratings yet
- 4 SignsDocument3 pages4 SignsJonathan GonzalezNo ratings yet
- Lit and PhilisophyDocument22 pagesLit and PhilisophyMJoyce Dela CruzNo ratings yet
- SDN PCo 03Document23 pagesSDN PCo 03UwedaNo ratings yet
- Daguyen, Shaira May K. 12-St. Agnes Trends, Networks, and Critical Thinking in The 21St Century Activity #3Document1 pageDaguyen, Shaira May K. 12-St. Agnes Trends, Networks, and Critical Thinking in The 21St Century Activity #3Aubrae Frances Engngeg BannawiNo ratings yet
- Characteristics of Transportation As A ProductDocument10 pagesCharacteristics of Transportation As A ProductSushmita Hatulan50% (2)