Download as pdf or txt
You might also like
- Breedlove SM Watson NV The Minds Machine Foundation of BrainDocument602 pagesBreedlove SM Watson NV The Minds Machine Foundation of BrainElena Chebotayeva100% (2)
- Anatomy and Physiology of BloodDocument9 pagesAnatomy and Physiology of BloodLharra Cagulada-PostranoNo ratings yet
- Anatomy and Physiology Case Study 2bDocument4 pagesAnatomy and Physiology Case Study 2bDylle Lorenzo ClaudioNo ratings yet
- Anatomy and Physiology of LeukemiaDocument7 pagesAnatomy and Physiology of LeukemiaJamaica Aurelio75% (4)
- Chastity Before Marriage - A Fresh Perspective PDFDocument18 pagesChastity Before Marriage - A Fresh Perspective PDFJeriz Marie GamboaNo ratings yet
- Megacode Testing Checklist: Scenarios 1/3/8 Bradycardia Pulseless VT PEA PCACDocument6 pagesMegacode Testing Checklist: Scenarios 1/3/8 Bradycardia Pulseless VT PEA PCACberril_fannyNo ratings yet
- Gjaku Dhe HemopoezaDocument56 pagesGjaku Dhe HemopoezaGent'No ratings yet
- Module 2 Blood and Bloostain Analysis Forensic 3Document29 pagesModule 2 Blood and Bloostain Analysis Forensic 3nikke requeNo ratings yet
- Tugas Anatomi IntegumenDocument26 pagesTugas Anatomi IntegumenLailia nur rachmaNo ratings yet
- 1378968920.48619-Blood - PDF Lab: March 2019Document19 pages1378968920.48619-Blood - PDF Lab: March 2019don aNo ratings yet
- Blood and Clotting FactorsDocument34 pagesBlood and Clotting FactorsAnand KasiNo ratings yet
- Anaphy Midterm Notes (The Blood Summary)Document4 pagesAnaphy Midterm Notes (The Blood Summary)Zosia Sage100% (1)
- Cardio Vascular SystemDocument18 pagesCardio Vascular SystemjeniferNo ratings yet
- Microsoft Word - The BloodDocument16 pagesMicrosoft Word - The BloodMelanie Fleckner0% (3)
- BloodDocument7 pagesBloodNaanmatha PuspanathanNo ratings yet
- Aspek Biokimiawi HematologiDocument72 pagesAspek Biokimiawi HematologiExcelsis Deo SombolinggiNo ratings yet
- Biochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Document21 pagesBiochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Pedro SilvaNo ratings yet
- Biochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Document28 pagesBiochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Pedro SilvaNo ratings yet
- Biochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Document26 pagesBiochemistry of Diseases Dr. Jishnunil Chakraborty: Analytical Clinical Biochemistry (Sec-A2)Pedro SilvaNo ratings yet
- Blood and Lymph: FunctionsDocument8 pagesBlood and Lymph: Functionsmadalina popaNo ratings yet
- Blood: Red Blood Cells (Erythrocytes)Document14 pagesBlood: Red Blood Cells (Erythrocytes)Pari JanNo ratings yet
- Components of BloodDocument10 pagesComponents of BloodDJ GAMARINo ratings yet
- A. Functions of Blood SystemDocument7 pagesA. Functions of Blood SystemPrant ChikyNo ratings yet
- Anatomy and Physiology of BloodDocument10 pagesAnatomy and Physiology of Bloodaoi_rachelle100% (1)
- Anatomy and Physiology of Blood and Cardiovascular SystemDocument6 pagesAnatomy and Physiology of Blood and Cardiovascular SystemCharise LigoresNo ratings yet
- Human Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions ManualDocument9 pagesHuman Physiology From Cells To Systems 8th Edition Lauralee Sherwood Solutions Manuallagantrongcnswd100% (33)
- Blood Overview and IntroductionDocument8 pagesBlood Overview and IntroductionKathleen Joy Costales MagtanongNo ratings yet
- Cito KrviDocument9 pagesCito KrviStrahinja SkoboNo ratings yet
- Blood - WikipediaDocument106 pagesBlood - WikipediaZoha AliNo ratings yet
- BloodDocument19 pagesBloodToday NewsNo ratings yet
- Blood Overview and IntroductionDocument8 pagesBlood Overview and IntroductionWai KikiNo ratings yet
- Blood and Blood Vessels Module 3Document16 pagesBlood and Blood Vessels Module 3Jenevieve Tungcul SimanganNo ratings yet
- Heamatology 1Document57 pagesHeamatology 1Victoria &BeautifulPeopleNo ratings yet
- Blood@Wikipedia Org enDocument18 pagesBlood@Wikipedia Org enHelmiNo ratings yet
- Blood Is A SpecializedDocument15 pagesBlood Is A SpecializedParthiv DaveNo ratings yet
- Blood: Figure 1: Shows Separated Blood Layers by CentrifugeDocument7 pagesBlood: Figure 1: Shows Separated Blood Layers by Centrifugeحسين مهديNo ratings yet
- Blood: Physiology and Circulation (Edited by Kara Rogers) : ReferenceDocument3 pagesBlood: Physiology and Circulation (Edited by Kara Rogers) : ReferenceJossefa AlmanitaNo ratings yet
- BloodDocument16 pagesBloodMohsin Bhat100% (1)
- BloodDocument4 pagesBloodatikarahmansuba123No ratings yet
- Transport System I - UnlockedDocument21 pagesTransport System I - Unlockeddannieycandiey44No ratings yet
- LomhematnotesDocument31 pagesLomhematnotesLibi FarrellNo ratings yet
- Functions of The Circulatory SystemDocument10 pagesFunctions of The Circulatory SystemAb AbNo ratings yet
- (Abastecimiento) (Tejidos) : Removal of Waste Such As Carbon Dioxide, Urea, and Lactic AcidDocument3 pages(Abastecimiento) (Tejidos) : Removal of Waste Such As Carbon Dioxide, Urea, and Lactic AcidMelissa TorresNo ratings yet
- Blood ReviewDocument35 pagesBlood ReviewCharli ParachinniNo ratings yet
- Ematology: MLT Department 2 - Stage Theoretical Part/ Lecture No. 1 by Dilgeer Ahmed KalariDocument24 pagesEmatology: MLT Department 2 - Stage Theoretical Part/ Lecture No. 1 by Dilgeer Ahmed Kalariphotocopy beshaNo ratings yet
- Leukemia Case StudyDocument10 pagesLeukemia Case StudyQueennieMarelRamos100% (1)
- Blood: From Wikipedia, The Free Encyclopedia Jump To:, For Other Uses, SeeDocument26 pagesBlood: From Wikipedia, The Free Encyclopedia Jump To:, For Other Uses, Seestev_brown89No ratings yet
- Blood FinalDocument14 pagesBlood FinalAnkit PattnayakNo ratings yet
- Citologija KrviDocument9 pagesCitologija KrviStrahinja SkoboNo ratings yet
- Blood: Mohamed Serar Yusuf Department of HistologyDocument42 pagesBlood: Mohamed Serar Yusuf Department of HistologyAboubakar Moalim Mahad moh'dNo ratings yet
- Text For Reding BloodDocument39 pagesText For Reding BloodFjhvbhhNo ratings yet
- Composition of BloodDocument5 pagesComposition of BloodMeghaNo ratings yet
- Lab 25 BloodDocument23 pagesLab 25 Bloodrhycelayon304No ratings yet
- B & C, Vol-11 PDFDocument4 pagesB & C, Vol-11 PDFKiran RmvNo ratings yet
- BCH 408Document13 pagesBCH 408ezekielolami01No ratings yet
- Chapter 17 Anatomy and Physiology NotesDocument10 pagesChapter 17 Anatomy and Physiology NotesZachary WatsonNo ratings yet
- Hematology SystemDocument15 pagesHematology SystemayuNo ratings yet
- Blood Anatomy and PhysiologyDocument4 pagesBlood Anatomy and PhysiologyRao Asad MubeenNo ratings yet
- Body Fluids and CirculationDocument5 pagesBody Fluids and CirculationSheehan MathurNo ratings yet
- Blood Anatomy and PhysiologyDocument23 pagesBlood Anatomy and PhysiologyNatukunda DianahNo ratings yet
- BloodDocument38 pagesBloodchukwukerechimezirimNo ratings yet
- GRP 5 Hematopoietic System ImlanDocument49 pagesGRP 5 Hematopoietic System ImlanSharie Grace ImlanNo ratings yet
- Office of The Secretary: FirstDocument12 pagesOffice of The Secretary: FirstJeriz Marie GamboaNo ratings yet
- Peptic Ulcer Disease: by Ntr/Jevr/Djs/AjisDocument14 pagesPeptic Ulcer Disease: by Ntr/Jevr/Djs/AjisJeriz Marie GamboaNo ratings yet
- Hepatocellular Carcinoma (HCC)Document37 pagesHepatocellular Carcinoma (HCC)Jeriz Marie GamboaNo ratings yet
- International Conference On Physical Activity and Obesity in Children: Summary Statement and RecommendationsDocument18 pagesInternational Conference On Physical Activity and Obesity in Children: Summary Statement and RecommendationsJeriz Marie GamboaNo ratings yet
- The Human Body: An Orientation: Chapter 1Document88 pagesThe Human Body: An Orientation: Chapter 1Jeriz Marie GamboaNo ratings yet
- Central Nervous System Infection in The PediatricDocument5 pagesCentral Nervous System Infection in The PediatricJeriz Marie GamboaNo ratings yet
- USTFMS ERB F01 Application FormDocument2 pagesUSTFMS ERB F01 Application FormJeriz Marie GamboaNo ratings yet
- Dating and CourtshipDocument12 pagesDating and CourtshipJeriz Marie GamboaNo ratings yet
- Source For EcolecDocument35 pagesSource For EcolecJeriz Marie GamboaNo ratings yet
- Make Health Your "New High" Life Not DrugsDocument1 pageMake Health Your "New High" Life Not DrugsJeriz Marie GamboaNo ratings yet
- Mitosis and Meiosis: Higher BiologyDocument16 pagesMitosis and Meiosis: Higher BiologyJeriz Marie GamboaNo ratings yet
- Xoxo GossipDocument2 pagesXoxo GossipJeriz Marie GamboaNo ratings yet
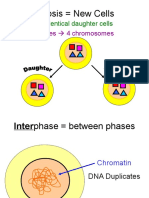
- Mitosis New Cells: Results in 2 Identical Daughter CellsDocument26 pagesMitosis New Cells: Results in 2 Identical Daughter CellsJeriz Marie GamboaNo ratings yet
- Human Genetic VariationDocument35 pagesHuman Genetic VariationJeriz Marie GamboaNo ratings yet
- Environmental and Experimental Botany: R. Cruz de Carvalho, A. Bernardes Da Silva, C. Branquinho, J. Marques Da SilvaDocument5 pagesEnvironmental and Experimental Botany: R. Cruz de Carvalho, A. Bernardes Da Silva, C. Branquinho, J. Marques Da SilvaJeriz Marie GamboaNo ratings yet
- Lesson 26 Communication Services: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionDocument36 pagesLesson 26 Communication Services: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionJeriz Marie GamboaNo ratings yet
- Lesson 27 Communications and Collaboration: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionDocument39 pagesLesson 27 Communications and Collaboration: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionJeriz Marie GamboaNo ratings yet
- Ekonomiks: Maxine Yosobelle P. Cruz 10-Righteousness Ms - SardonDocument5 pagesEkonomiks: Maxine Yosobelle P. Cruz 10-Righteousness Ms - SardonJeriz Marie Gamboa0% (1)
- Lesson 28 Using The Internet and The World Wide Web: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionDocument39 pagesLesson 28 Using The Internet and The World Wide Web: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionJeriz Marie GamboaNo ratings yet
- Lesson 25 Network Fundamentals: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionDocument45 pagesLesson 25 Network Fundamentals: Computer Literacy Basics: A Comprehensive Guide To IC, 5 EditionJeriz Marie GamboaNo ratings yet
- NCM 112 - Drug StudyDocument35 pagesNCM 112 - Drug StudyZoe WsetNo ratings yet
- IV Fluid Management: Islam Awni Abu SamraDocument41 pagesIV Fluid Management: Islam Awni Abu SamraIslam AwniNo ratings yet
- Shock AssessmentDocument36 pagesShock AssessmentAlyssandra LucenoNo ratings yet
- MNT For Metabolic StressDocument107 pagesMNT For Metabolic StressMeyar Y. Abu AsadNo ratings yet
- Rollplay InggrisDocument4 pagesRollplay InggrisIrmaNo ratings yet
- Breath of Fire - KundaliniDocument2 pagesBreath of Fire - KundaliniErica Yang100% (1)
- Answer KeyDocument5 pagesAnswer KeyJhoey Riparip PapaNo ratings yet
- MCQ FMS 3 (2016) With Answers-1Document17 pagesMCQ FMS 3 (2016) With Answers-1stella pangestikaNo ratings yet
- ATPL 040.human Factor NotesDocument22 pagesATPL 040.human Factor NotesAbu BrahimNo ratings yet
- NurseLabs QnA#1Document41 pagesNurseLabs QnA#1Jill NoelleNo ratings yet
- Liver Cirrhosis: DR - Aryadevi A SDocument89 pagesLiver Cirrhosis: DR - Aryadevi A SMahesh PethkarNo ratings yet
- Cardiac ArrestDocument30 pagesCardiac ArrestTommy NainggolanNo ratings yet
- Laboratory Worksheet 12 Heart Structure and FunctionDocument4 pagesLaboratory Worksheet 12 Heart Structure and FunctionAndrea SaldivarNo ratings yet
- Warm-Up & CooldownDocument2 pagesWarm-Up & CooldownJacquelineMarieAndresNo ratings yet
- BedwettingDocument3 pagesBedwettingaanya khajuriaNo ratings yet
- CopdDocument5 pagesCopdapi-719803017No ratings yet
- Review:: Although Cardiac Troponin (CTN) Is ADocument15 pagesReview:: Although Cardiac Troponin (CTN) Is ACristina DobrinNo ratings yet
- Concept Map KawasakiDocument6 pagesConcept Map KawasakiBrent John Trasmonte100% (1)
- Homework-Alkalosis and AcidosisDocument2 pagesHomework-Alkalosis and AcidosisPatricia LacanlaleNo ratings yet
- 1st Periodic Exam g9Document3 pages1st Periodic Exam g9Vincent Franciz T. FernandezNo ratings yet
- Anatomy and Pathophysiology of AnemiaDocument9 pagesAnatomy and Pathophysiology of AnemiaRaja SharmaNo ratings yet
- Biokimia 2 GLIKOLISISDocument18 pagesBiokimia 2 GLIKOLISISnamharNo ratings yet
- Fundamentals of Anatomy, Physiology in Sports-1Document15 pagesFundamentals of Anatomy, Physiology in Sports-1avik mukherjeeNo ratings yet
- Cardiovascular Practice Questions Answers and RationalesFALL2019-1Document11 pagesCardiovascular Practice Questions Answers and RationalesFALL2019-1titilayo loveNo ratings yet
- English (Respiratory System)Document16 pagesEnglish (Respiratory System)ChitraNo ratings yet
- Psychophysiological Study of Out-of-the-Body Experiences in A Selected SubjectDocument7 pagesPsychophysiological Study of Out-of-the-Body Experiences in A Selected SubjectIno MoxoNo ratings yet
- Hemodynamic Monitoring For NursesDocument29 pagesHemodynamic Monitoring For NursesMahalakshmi Prabhu100% (1)
- NCP CopdDocument6 pagesNCP Copdjoelette100% (1)