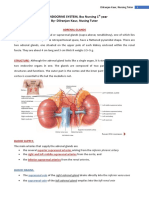
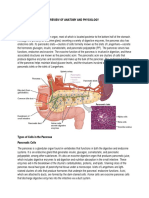
Download as pdf or txt
You might also like
- Extractable Base Neutrals and AcidsDocument15 pagesExtractable Base Neutrals and AcidsTrisyan VikyNo ratings yet
- Cell InjuryDocument118 pagesCell InjuryShanzayNo ratings yet
- Trace Elements and Porphyrins - JBDocument7 pagesTrace Elements and Porphyrins - JBVinNo ratings yet
- Cell Injury & AdaptationDocument6 pagesCell Injury & AdaptationVita Dwi PayantiNo ratings yet
- Lab Report On Gene Cloning and Vector ExpressionDocument11 pagesLab Report On Gene Cloning and Vector Expressionselina_kolls100% (2)
- Recombinant DNA TechnologyDocument108 pagesRecombinant DNA TechnologyselowestNo ratings yet
- Protein Expression HandbookDocument118 pagesProtein Expression HandbookLuis Arístides Torres SánchezNo ratings yet
- Role of NF-Binthe Regulation of Immunity and Apoptosis: Kavitha BharathamDocument30 pagesRole of NF-Binthe Regulation of Immunity and Apoptosis: Kavitha BharathamArief Adi SaputroNo ratings yet
- Chronic Myelogenous Leukemia (CML) - Causes, Symptoms, TreatmentDocument10 pagesChronic Myelogenous Leukemia (CML) - Causes, Symptoms, Treatmentnurul auliaNo ratings yet
- Eicosanoids: (Prostaglandins, Thromboxanes, Leukotrienes)Document26 pagesEicosanoids: (Prostaglandins, Thromboxanes, Leukotrienes)BOsch VakilNo ratings yet
- The Complement SystemDocument24 pagesThe Complement Systemhkatniwala100% (1)
- Topic: Medicial Chemistry of HormonesDocument18 pagesTopic: Medicial Chemistry of HormonesHaris AliNo ratings yet
- Chronic Obstructive Pulmonary DiseaseDocument2 pagesChronic Obstructive Pulmonary DiseaseShrestha AnjivNo ratings yet
- Systemic Lupus Erythematosus Lecture Salah Abdel BakyDocument8 pagesSystemic Lupus Erythematosus Lecture Salah Abdel Bakyahmed gabrNo ratings yet
- Patho Final Study GuideDocument55 pagesPatho Final Study GuideBritNo ratings yet
- Pharmacodynamics3 161031154812Document43 pagesPharmacodynamics3 161031154812Adeel Khan100% (1)
- Endo 3 Notes PDFDocument9 pagesEndo 3 Notes PDFDilNo ratings yet
- Po 65264 LC Ms Quant Analysis Steroids Serum Asms2018 Po65264 enDocument1 pagePo 65264 LC Ms Quant Analysis Steroids Serum Asms2018 Po65264 enMôùhamed BnsNo ratings yet
- SaveBooklet BookletDocument120 pagesSaveBooklet BookletRoscelie KhoNo ratings yet
- Complement SystemDocument6 pagesComplement SystemJimit GandhiNo ratings yet
- 2021 Host Reponse To InfectionDocument38 pages2021 Host Reponse To Infectionmaggie100% (1)
- Pa Tho Physiology of ParkinsonDocument2 pagesPa Tho Physiology of Parkinsoncuriosity_killsNo ratings yet
- Hormone Feedback LoopsDocument6 pagesHormone Feedback LoopsSylheti BabaNo ratings yet
- Pku AlkDocument33 pagesPku AlkSharf WangNo ratings yet
- Mechanisms of Cell InjuryDocument21 pagesMechanisms of Cell Injuryبراءة أحمد السلاماتNo ratings yet
- Review of Anatomy and PhysiologyDocument7 pagesReview of Anatomy and PhysiologyKyla CalzadoNo ratings yet
- Autocoid PharmacologyDocument29 pagesAutocoid PharmacologyLyadelou Fortu100% (1)
- Vii. Laboratory Results: Hemoglobin 129 MG/DL 140-160 MG/DL LowDocument2 pagesVii. Laboratory Results: Hemoglobin 129 MG/DL 140-160 MG/DL LowAdrian Dela CruzNo ratings yet
- Thyroid Hormones and AntiDocument2 pagesThyroid Hormones and AntiNishaHasanNo ratings yet
- Idiopathic Pulmonary Fibrosis: Optimizing The Diagnosis and Multi-Disciplinary Decision MakingDocument4 pagesIdiopathic Pulmonary Fibrosis: Optimizing The Diagnosis and Multi-Disciplinary Decision MakingAnna LiachenkoNo ratings yet
- G-protein-Coupled ReceptorsDocument24 pagesG-protein-Coupled ReceptorsNaimi Amalia hatimahNo ratings yet
- Histamine 5-ht Angiotensin Kinin Endothelins Eicosanoids Interleukins TNF Interferones Growth FCDocument57 pagesHistamine 5-ht Angiotensin Kinin Endothelins Eicosanoids Interleukins TNF Interferones Growth FCFaisal 'arifNo ratings yet
- Classification of The Epilepsies: Purpose: For Clinical DiagnosisDocument25 pagesClassification of The Epilepsies: Purpose: For Clinical Diagnosisayu rifqiNo ratings yet
- Topic: Cytokine and Their Roles in The Immune Regulation: Md. Jiyaul MustafaDocument21 pagesTopic: Cytokine and Their Roles in The Immune Regulation: Md. Jiyaul MustafaAisha Siddiq100% (1)
- 2nd Peroxisome Metabolism 20760414Document56 pages2nd Peroxisome Metabolism 20760414Rawbeena RamtelNo ratings yet
- The Resistance Offered by The Host Towards Injury Caused by Microorganisms and Their Products or The State of Protection From Infectious DiseaseDocument21 pagesThe Resistance Offered by The Host Towards Injury Caused by Microorganisms and Their Products or The State of Protection From Infectious DiseaseReshmiAkhilNo ratings yet
- Causes of Stroke PDFDocument16 pagesCauses of Stroke PDFEmmanuel AguilarNo ratings yet
- BIOL 243 Guided Practice 3 - Transport Flow Chart AssignmentDocument1 pageBIOL 243 Guided Practice 3 - Transport Flow Chart AssignmentReeseNo ratings yet
- Intro + Pharmacodynamics 2Document46 pagesIntro + Pharmacodynamics 2Dana E AbuqaudNo ratings yet
- Analyte Stability & Freeze-Thaw Information-1Document8 pagesAnalyte Stability & Freeze-Thaw Information-1Yusuf Indra SentosaNo ratings yet
- Classification of AnemiaDocument4 pagesClassification of AnemiaEna PaparićNo ratings yet
- Graves' Disease: Bella I. Putri 16-025Document29 pagesGraves' Disease: Bella I. Putri 16-025anon_414347743No ratings yet
- Widal TestDocument2 pagesWidal TestRavi AhirwarNo ratings yet
- Portal Vein Thrombus ChildrenDocument11 pagesPortal Vein Thrombus ChildrenViswas ChhapolaNo ratings yet
- Electroporation: Jac A NickoloffDocument3 pagesElectroporation: Jac A Nickoloffmanoj_rkl_07No ratings yet
- Steroids HormonesDocument15 pagesSteroids HormonesZain Ul Abdin Mughal100% (1)
- Pathophysiology of FeverDocument4 pagesPathophysiology of FeverEasha Jamil AbbasiNo ratings yet
- Apoptosis: Apo: Apart Ptosis: FallenDocument28 pagesApoptosis: Apo: Apart Ptosis: Fallengull naz100% (1)
- Glycogen Storage Disease, Type II (Pompe Disease) : BackgroundDocument7 pagesGlycogen Storage Disease, Type II (Pompe Disease) : Background201111000No ratings yet
- Morales J. The Innate Immune System in Health... Vol 2. 2022Document312 pagesMorales J. The Innate Immune System in Health... Vol 2. 2022Philip Rodríguez GarcíaNo ratings yet
- Basic Principles of Cell Injury and AdaptationDocument61 pagesBasic Principles of Cell Injury and Adaptationbesthachakrapani100% (5)
- Alzheimer's DiseaseDocument19 pagesAlzheimer's DiseaseMission JupiterNo ratings yet
- Crispr TechniqueDocument16 pagesCrispr TechniqueAmmar Abbas100% (1)
- Inborn Errors of MetabolismDocument4 pagesInborn Errors of Metabolismcurly perkyNo ratings yet
- 2 Cell Injury, Adaptation, and Cell DeathDocument6 pages2 Cell Injury, Adaptation, and Cell DeathcedonuliNo ratings yet
- Day 1 BacteriaDocument251 pagesDay 1 BacteriaLisa NetherlandNo ratings yet
- Walking Pneumonia, (Mycoplasma Pneumonia) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandWalking Pneumonia, (Mycoplasma Pneumonia) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- Gastrointestinal Diseases and Disorders Sourcebook, Fifth EditionFrom EverandGastrointestinal Diseases and Disorders Sourcebook, Fifth EditionNo ratings yet
- Hum. Reprod.-2006-García-Pérez-880-7Document8 pagesHum. Reprod.-2006-García-Pérez-880-7ANIS MURNIATINo ratings yet
- Vector Biology: Cloning and Expression Vectors: Group 3Document32 pagesVector Biology: Cloning and Expression Vectors: Group 3Akinade AdetolaNo ratings yet
- TFS ProteinresearchhandbookDocument126 pagesTFS ProteinresearchhandbookSupun Sandaru FernandoNo ratings yet
- Expression VectorDocument11 pagesExpression VectorSURBHI SINGHALNo ratings yet
- Expression Vectors PDFDocument2 pagesExpression Vectors PDFJennifer33% (3)
- List of PH.D Thesis at IvriDocument21 pagesList of PH.D Thesis at IvriMuguo Mouafo BriceNo ratings yet
- Plasmid pBR322Document38 pagesPlasmid pBR322Adriana MorenoNo ratings yet
- BIO374 Fall 2018 Recombinant DNA and GenomicsDocument30 pagesBIO374 Fall 2018 Recombinant DNA and GenomicsdanNo ratings yet
- Cloning VectorsDocument161 pagesCloning Vectorssubhash nayak100% (2)
- Production of Recombinant Therapeutic ProteinsDocument28 pagesProduction of Recombinant Therapeutic ProteinstasniyanNo ratings yet
- Manipulasi Dan Ekspresi GenDocument40 pagesManipulasi Dan Ekspresi GenamandaNo ratings yet
- Dino1 BlastnDocument217 pagesDino1 Blastnlionkonsti37No ratings yet
- Cloning and Expression - FLAG and 3xFLAG OverviewDocument3 pagesCloning and Expression - FLAG and 3xFLAG OverviewSigma-Aldrich100% (12)
- Q Paper MBT-CC301Document2 pagesQ Paper MBT-CC301SameerNo ratings yet
- ArcticExpress Competent Cells-Manual IDocument18 pagesArcticExpress Competent Cells-Manual ILarissa GoncalvesNo ratings yet
- Expression of Cloned Protein in Animal CellDocument4 pagesExpression of Cloned Protein in Animal CellSadiqa ForensicNo ratings yet
- Expression of Proteins in Animal Cell Culture System: RecombinantDocument26 pagesExpression of Proteins in Animal Cell Culture System: RecombinantSwateja MangleNo ratings yet
- Enhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator GenesDocument7 pagesEnhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator Geneszhaoyue12112001No ratings yet
- Bio 102 Practice Problems Recombinant DNA and BiotechnologyDocument7 pagesBio 102 Practice Problems Recombinant DNA and BiotechnologyI FNo ratings yet
- DNA Virus Vectors For Vaccine ProductionDocument12 pagesDNA Virus Vectors For Vaccine ProductionSAMANTHA DE JESUS RIVERO MONTEJONo ratings yet
- Insect Cell CultureDocument6 pagesInsect Cell CultureJakobsen2012No ratings yet
- Proteínaa BrazzeinDocument6 pagesProteínaa BrazzeinDiana TorresNo ratings yet
- Strategies To Maximize Heterologous Protein ExpressionDocument10 pagesStrategies To Maximize Heterologous Protein ExpressionElías Octavio Gómez MontesNo ratings yet
- Pichia Manual PDFDocument100 pagesPichia Manual PDFhana kimiNo ratings yet
- Donini, Marusic - 2019 - Current State-Of-The-Art in Plant-Based Antibody Production Systems PDFDocument12 pagesDonini, Marusic - 2019 - Current State-Of-The-Art in Plant-Based Antibody Production Systems PDFAndrei TatomirNo ratings yet
- Selection of rDNADocument31 pagesSelection of rDNADeepika KV100% (1)
- Cloning StrategiesDocument41 pagesCloning StrategiesKumar Aavula100% (2)
- En Ni NTA Superflow BioRobot HandbookDocument64 pagesEn Ni NTA Superflow BioRobot HandbookAdolfo CruzNo ratings yet