Download as pdf or txt
You might also like
- Annett Book 2-1Document201 pagesAnnett Book 2-1Benedetta Flebus100% (1)
- M Phase in An Animal CellDocument2 pagesM Phase in An Animal CellJohnNo ratings yet
- 10-11, Kinetics of Enzyme Catalyzed ReactionDocument34 pages10-11, Kinetics of Enzyme Catalyzed ReactionS. Ansary100% (1)
- Enzyme M&LDocument6 pagesEnzyme M&LNethra Sasikumar100% (1)
- Enzyme KineticsDocument18 pagesEnzyme KineticsRoyal MindNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- Enzyme and Acid - Base CatalysisDocument64 pagesEnzyme and Acid - Base Catalysisbinseung skzNo ratings yet
- Biology 3601 Biochemistry Enzyme Kinetics Laboratory BackgroundDocument5 pagesBiology 3601 Biochemistry Enzyme Kinetics Laboratory Backgroundkgeorges27No ratings yet
- Michaelis Menten Equation SummaryDocument7 pagesMichaelis Menten Equation SummaryBüşra GültekinNo ratings yet
- MIT Lecture 6 5.07 Biochemistry LectureDocument16 pagesMIT Lecture 6 5.07 Biochemistry LectureakiridoNo ratings yet
- BS 3Document60 pagesBS 3himanshu_agraNo ratings yet
- Michaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)Document2 pagesMichaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)fintastellaNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Michaelis Menten EquationDocument9 pagesMichaelis Menten Equationsadaf zaidiNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- LEC11 EnzKin 08Document12 pagesLEC11 EnzKin 08princesschemistNo ratings yet
- Enzyme NotesDocument18 pagesEnzyme NotesDBPNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- Lecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Document6 pagesLecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Louis FortunatoNo ratings yet
- Lecture Notes Enzyme 2 Enzyme Kinetics WebDocument29 pagesLecture Notes Enzyme 2 Enzyme Kinetics WebAldren RebaLdeNo ratings yet
- Lecture 12 - Enzyme Kinetics IDocument30 pagesLecture 12 - Enzyme Kinetics IThomas Jones50% (2)
- Solutions Chapter8Document25 pagesSolutions Chapter8Fábio RamalhoNo ratings yet
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Enzymes - Michael L. ShulerDocument23 pagesEnzymes - Michael L. ShulerAndrésFelipeQuirogaMirandaNo ratings yet
- Enzyme Kinetics: Medical Biochemistry, Lecture 24Document39 pagesEnzyme Kinetics: Medical Biochemistry, Lecture 24jeyankarunanithiNo ratings yet
- Lecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesDocument21 pagesLecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesAkash AroraNo ratings yet
- Enzyme Kinetics Practical For RecordingDocument22 pagesEnzyme Kinetics Practical For RecordingSanya chauhanNo ratings yet
- EnzomologyDocument26 pagesEnzomologyToga Brandon100% (1)
- Enzyme KineticsDocument23 pagesEnzyme Kinetics1DS19CH011 Jashwanth C RNo ratings yet
- MichaelisDocument4 pagesMichaelishamzaasghar233No ratings yet
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Michaelis Menten DerivedDocument4 pagesMichaelis Menten DerivedGOOGLI TECNo ratings yet
- Immobilized Enzyme ReactorsDocument33 pagesImmobilized Enzyme ReactorsRAKESH V RNo ratings yet
- Enzyme InhibitionDocument4 pagesEnzyme InhibitionNisha ManasNo ratings yet
- The Energy Charge of The Adenylate Pool As A Regulatory Parameter. Interaction With Feedback Modifiers"Document5 pagesThe Energy Charge of The Adenylate Pool As A Regulatory Parameter. Interaction With Feedback Modifiers"rahul vivekNo ratings yet
- Lecture 3 Notes - EnzymologyDocument9 pagesLecture 3 Notes - EnzymologyYi Gong100% (3)
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Document16 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Ken M'voulaboloNo ratings yet
- Enzyme Catalysis PDFDocument17 pagesEnzyme Catalysis PDFuzzal ahmedNo ratings yet
- Dokumen - Tips - Enzymes Problem SetDocument22 pagesDokumen - Tips - Enzymes Problem SetDawlat SlamaNo ratings yet
- MIT5 07SCF13 Lec7 8Document19 pagesMIT5 07SCF13 Lec7 8Ruben RodriguezNo ratings yet
- Enzyme2 2012Document45 pagesEnzyme2 2012Surya Prakash KabiNo ratings yet
- Factors AffectingDocument11 pagesFactors Affectingshekinah656No ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsalicjadzNo ratings yet
- Teknik ReaksiDocument21 pagesTeknik ReaksiAhmadi FachryNo ratings yet
- CB Chapter3Document30 pagesCB Chapter3Leidy IracemaNo ratings yet
- Dr. Nazir Ahmad: Nehal BilalDocument15 pagesDr. Nazir Ahmad: Nehal BilalAayat MughalNo ratings yet
- Determination of EnzymesDocument8 pagesDetermination of EnzymesUsman GhaniNo ratings yet
- 1 EnzymesDocument30 pages1 EnzymesRohit pansareNo ratings yet
- Michaelis-Menten KineticsDocument6 pagesMichaelis-Menten Kineticscharanmann9165No ratings yet
- 2 Bioprocess Engineering CH 2Document54 pages2 Bioprocess Engineering CH 2Fasil ManNo ratings yet
- Lecture 17: Enzyme Kinetics: Biochemistry I Oct 8, 2007Document2 pagesLecture 17: Enzyme Kinetics: Biochemistry I Oct 8, 2007zdfgarsgNo ratings yet
- Enzymes Kinetics & RegulationDocument41 pagesEnzymes Kinetics & RegulationConorTankGorbatsjovNo ratings yet
- Chem 40 Enzyme KineticsDocument85 pagesChem 40 Enzyme KineticsJustine Grace Mariano100% (1)
- 2 Biochemical Engineering CH 2Document54 pages2 Biochemical Engineering CH 2Fasil ManNo ratings yet
- En ZymologyDocument24 pagesEn Zymologywyclife akong'oNo ratings yet
- Introduction To Enzyme KineticsDocument3 pagesIntroduction To Enzyme KineticsSilvia Paulina Monrroy CifuentesNo ratings yet
- Michaelis MentenKineticsandBriggs HaldaneKinetics - HTMLDocument2 pagesMichaelis MentenKineticsandBriggs HaldaneKinetics - HTMLCrestore Lex Tapia CapiñaNo ratings yet
- IIPE Class 4 24.03Document41 pagesIIPE Class 4 24.03bharatNo ratings yet
- Lecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003Document4 pagesLecture 15: Enzyme Inhibition: Biochemistry I Fall Term, 2003archerofthestarsNo ratings yet
- Theory and Applications of the Empirical Valence Bond Approach: From Physical Chemistry to Chemical BiologyFrom EverandTheory and Applications of the Empirical Valence Bond Approach: From Physical Chemistry to Chemical BiologyFernanda DuarteNo ratings yet
- X Ray Diffraction 1057Document5 pagesX Ray Diffraction 1057JohnNo ratings yet
- Open Educational Resources Textbook ListDocument35 pagesOpen Educational Resources Textbook ListJohn100% (1)
- XML For Software Engineers: Tutorial OutlineDocument47 pagesXML For Software Engineers: Tutorial OutlineJohnNo ratings yet
- Exponential Notation and Physical QuantitiesDocument4 pagesExponential Notation and Physical QuantitiesJohnNo ratings yet
- Plant Tissue SystemsDocument2 pagesPlant Tissue SystemsJohn0% (1)
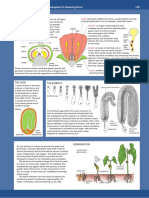
- Flower DevelopmentDocument1 pageFlower DevelopmentJohnNo ratings yet
- PolymerizationDocument2 pagesPolymerizationJohnNo ratings yet
- Actin Filaments and MicrotubulesDocument2 pagesActin Filaments and MicrotubulesJohnNo ratings yet
- Actin Filaments: 968 PANEL 16-1: The Three Major Types of Protein Filaments That Form The CytoskeletonDocument1 pageActin Filaments: 968 PANEL 16-1: The Three Major Types of Protein Filaments That Form The CytoskeletonJohnNo ratings yet
- Signal Sequences and Protein TranslocationDocument1 pageSignal Sequences and Protein TranslocationJohnNo ratings yet
- Genes and PhenotypesDocument2 pagesGenes and PhenotypesJohnNo ratings yet
- Sources of Intracellular OsmolarityDocument1 pageSources of Intracellular OsmolarityJohnNo ratings yet
- Hydrogen BondsDocument2 pagesHydrogen BondsJohnNo ratings yet
- Depicting A Small ProteinDocument2 pagesDepicting A Small ProteinJohnNo ratings yet
- Citric Acid CycleDocument2 pagesCitric Acid CycleJohnNo ratings yet
- Weak Chemical BondsDocument2 pagesWeak Chemical BondsJohnNo ratings yet
- Amino AcidsDocument2 pagesAmino AcidsJohnNo ratings yet
- Common Fatty AcidsDocument2 pagesCommon Fatty AcidsJohnNo ratings yet
- Use of Pyroxenite in Place of DolomiteDocument4 pagesUse of Pyroxenite in Place of DolomiteDhirender KapoorNo ratings yet
- Material Data Sheet For NH25-35MICDocument2 pagesMaterial Data Sheet For NH25-35MICJean-Noël LerouxNo ratings yet
- Product - Info G 2 B Annealing Furnace GLO ENDocument12 pagesProduct - Info G 2 B Annealing Furnace GLO ENBenhazen Lynn DeirdreNo ratings yet
- Chlorine Versus Twinoxide: The Answer To The Traditional ProblemDocument2 pagesChlorine Versus Twinoxide: The Answer To The Traditional ProblemTeknik MarinaNo ratings yet
- COM 08 11328asdfdsDocument9 pagesCOM 08 11328asdfdsVictor NgNo ratings yet
- Buffer Systems in The Body: Protein Buffers in Blood Plasma and CellsDocument11 pagesBuffer Systems in The Body: Protein Buffers in Blood Plasma and CellsK Jayakumar KandasamyNo ratings yet
- LESSON PLAN IN PHYSICAL SCIENCE (Sample Only)Document2 pagesLESSON PLAN IN PHYSICAL SCIENCE (Sample Only)Charmine AbuanNo ratings yet
- EM2210 Manual enDocument18 pagesEM2210 Manual enPeter SuNo ratings yet
- Pressure Drop and Velocity Simulations in Non-Stochastic Lattice Structure For Filter Applications Fabricated Using Additive ManufacturingDocument8 pagesPressure Drop and Velocity Simulations in Non-Stochastic Lattice Structure For Filter Applications Fabricated Using Additive ManufacturingEduardo Leal SalgadoNo ratings yet
- 0620 s05 Ms 3Document7 pages0620 s05 Ms 3Varun PanickerNo ratings yet
- Lecture05 Physical LayercontdDocument23 pagesLecture05 Physical LayercontdRehan BasharatNo ratings yet
- Mark Scheme (Results) January 2020: Pearson Edexcel International GCSE in Physics (4PH1) Paper 1PDocument17 pagesMark Scheme (Results) January 2020: Pearson Edexcel International GCSE in Physics (4PH1) Paper 1PNairit60% (5)
- Test 1 Review-SolutionsDocument11 pagesTest 1 Review-SolutionsdavidchoikekekekNo ratings yet
- Acids and Bases Weebly FileDocument5 pagesAcids and Bases Weebly Fileapi-240469044No ratings yet
- Naphthenic Acid Analysis Tan - Nan - Nat-Ms: COQG Meeting September 29, 2005Document14 pagesNaphthenic Acid Analysis Tan - Nan - Nat-Ms: COQG Meeting September 29, 2005Indra NainggolanNo ratings yet
- Kinetic Theory of Gases Powerpoint 2023Document24 pagesKinetic Theory of Gases Powerpoint 2023Nickolai AntoineNo ratings yet
- Earth Science Summative TestDocument4 pagesEarth Science Summative TestRoselle VictoriaNo ratings yet
- Eftekhari Sis2013Document86 pagesEftekhari Sis2013Luis MartinezNo ratings yet
- PC2132 Syllabus NUSDocument7 pagesPC2132 Syllabus NUSChris KingNo ratings yet
- Iron Powder MetallurgyDocument388 pagesIron Powder MetallurgyUmer MuzammilNo ratings yet
- Measuring Your Carbon Footprint PowerPointDocument16 pagesMeasuring Your Carbon Footprint PowerPointpremquilonNo ratings yet
- GHS Vs MSDS FormatsDocument5 pagesGHS Vs MSDS Formatse1717y8653No ratings yet
- 55NO3LTADocument14 pages55NO3LTASantiago DiosdadoNo ratings yet
- ImmunoelectrophoresisDocument3 pagesImmunoelectrophoresiskashishNo ratings yet
- BIOL1408 Introductory Biology Name Lab Unit 6/7: Diffusion & Osmosis Date Dr. Flo OxleyDocument10 pagesBIOL1408 Introductory Biology Name Lab Unit 6/7: Diffusion & Osmosis Date Dr. Flo OxleyJavier E. Dubon0% (1)
- Fenske Underwood Gilliland (FUG)Document8 pagesFenske Underwood Gilliland (FUG)HariKrishnaBushiNo ratings yet
- Aci sp-221-2004Document978 pagesAci sp-221-2004Abigael Valles RamirezNo ratings yet
- 1.2.1 PF Filter PlatesDocument52 pages1.2.1 PF Filter PlatesCristian Pulgar Fabres100% (2)
- Safety Data Sheet: NPK/NP/NK ( 70% AN) : Non-HazardousDocument16 pagesSafety Data Sheet: NPK/NP/NK ( 70% AN) : Non-HazardousMahmoud MahmoudmNo ratings yet