Download as pdf or txt
You might also like
- Novel Technique For Management of Bartholin Gland CystsnDocument3 pagesNovel Technique For Management of Bartholin Gland CystsnelenNo ratings yet
- How To Have Efficient SexDocument85 pagesHow To Have Efficient SexShivam Agrawal100% (2)
- Role of Leptin in ReproductionDocument10 pagesRole of Leptin in ReproductionCromironNo ratings yet
- Nutrition and Female FertilityDocument6 pagesNutrition and Female FertilityNumber MathematicianNo ratings yet
- Ovarian ReserverDocument35 pagesOvarian Reserverpramod singhNo ratings yet
- Childs Et Al., 2021 INGDocument56 pagesChilds Et Al., 2021 INGXochitemiqui YoacihualtNo ratings yet
- The Ovarian ReserveDocument19 pagesThe Ovarian Reservepramod singhNo ratings yet
- FSHR and LHR in Granulosa CellsDocument13 pagesFSHR and LHR in Granulosa CellsBhupathi SukeshNo ratings yet
- Nutrition and Female FertilityDocument6 pagesNutrition and Female FertilityFabiana Mejia MendezNo ratings yet
- Scaramuzzi 2011 PDFDocument24 pagesScaramuzzi 2011 PDFKarol Mariam VelázquezNo ratings yet
- Nutritional and Environmental Effects OnDocument11 pagesNutritional and Environmental Effects OnHassen AhmedNo ratings yet
- Studying The Role of Leptinand Adiponectin Hormones and Their Relation With Some Reproductive Hormones in Breastfeeding WomenDocument9 pagesStudying The Role of Leptinand Adiponectin Hormones and Their Relation With Some Reproductive Hormones in Breastfeeding WomenadraaNo ratings yet
- Abstract Placenta AustráliaDocument111 pagesAbstract Placenta AustráliaMilena MoraesNo ratings yet
- Nutrition and Reproduction 1996 RobinsonDocument10 pagesNutrition and Reproduction 1996 RobinsonCanuto MunozNo ratings yet
- Meyer Et Al. 2004Document15 pagesMeyer Et Al. 2004Aline Freitas de MeloNo ratings yet
- 5, Kisspeptin and PubertyDocument7 pages5, Kisspeptin and PubertyImy Imanلهثغ٨بع٨لعقحهحهخفتلحت٨ •No ratings yet
- Placenta 2Document21 pagesPlacenta 2Nurba Enda Karina NasutionNo ratings yet
- 1479 6805 Joe 21 0449Document12 pages1479 6805 Joe 21 0449Salim MichaelNo ratings yet
- Farooqi & O'Rahilly, 2009 - Leptin, A Pivotal Regulator of Human Energy HomeostasisDocument5 pagesFarooqi & O'Rahilly, 2009 - Leptin, A Pivotal Regulator of Human Energy HomeostasisMara ParamoNo ratings yet
- Perspective: Leptin's Physiologic Role: Does The Emperor of Energy Balance Have No Clothes?Document3 pagesPerspective: Leptin's Physiologic Role: Does The Emperor of Energy Balance Have No Clothes?ShuiNo ratings yet
- (17417899 - Reproduction) C-Type Natriuretic Peptide Enhances Mouse Preantral Follicle Growth-1Document11 pages(17417899 - Reproduction) C-Type Natriuretic Peptide Enhances Mouse Preantral Follicle Growth-1Dr. Eser AĞARNo ratings yet
- 2024 04 23 - PaperDocument20 pages2024 04 23 - PaperdrmanashreesankheNo ratings yet
- Jurnal Katak PDFDocument5 pagesJurnal Katak PDF31122007No ratings yet
- Fleming 2010Document4 pagesFleming 2010Ahmed GhanimNo ratings yet
- Hum. Reprod. Update 2009 Gellersen 119 38Document20 pagesHum. Reprod. Update 2009 Gellersen 119 38ayluyNo ratings yet
- New Insights Into The Role of LH in Early Ovarian Follicular Growth. A Possible Tool To Optimize Follicular Recruitment. La Marca 2023Document11 pagesNew Insights Into The Role of LH in Early Ovarian Follicular Growth. A Possible Tool To Optimize Follicular Recruitment. La Marca 2023Antonio JiménezNo ratings yet
- ViewtaegeDocument2 pagesViewtaegemihrullah.azimi1No ratings yet
- FTO PaperDocument9 pagesFTO PapervelshteinNo ratings yet
- Stress and The Female Reproductive System: S.N. Kalantaridou, A. Makrigiannakis, E. Zoumakis, G.P. ChrousosDocument8 pagesStress and The Female Reproductive System: S.N. Kalantaridou, A. Makrigiannakis, E. Zoumakis, G.P. ChrousosIrul AnwarNo ratings yet
- Lonergan 2016Document9 pagesLonergan 2016Resita ReiitaNo ratings yet
- (17417899 - Reproduction) A Link Between Notch and Progesterone Maintains The Functionality of The Rat Corpus LuteumDocument10 pages(17417899 - Reproduction) A Link Between Notch and Progesterone Maintains The Functionality of The Rat Corpus Luteumتقوى اللهNo ratings yet
- E443 FullDocument14 pagesE443 FullMarianaafiatiNo ratings yet
- J.theriogenology.2016.03.032 MetodologiaDocument6 pagesJ.theriogenology.2016.03.032 MetodologiaALEXA DANNAE MERLOS DIAZNo ratings yet
- Aerts Et Al., 2018 - Evaluation of A Role For NPY and NPY2R in The Pathogenesis of Obestiy by Mutation and Copy Number Variation Analysis in Obese Children and AdolescentsDocument10 pagesAerts Et Al., 2018 - Evaluation of A Role For NPY and NPY2R in The Pathogenesis of Obestiy by Mutation and Copy Number Variation Analysis in Obese Children and AdolescentsLautaro AndradeNo ratings yet
- Extracellular Nucleic Acids in Maternal Circulation As Potential Biomarkers For Placental InsufficiencyDocument12 pagesExtracellular Nucleic Acids in Maternal Circulation As Potential Biomarkers For Placental Insufficiencymaccio88No ratings yet
- Antibiotic Exposure Promotes Fat GainDocument3 pagesAntibiotic Exposure Promotes Fat GainCarolina Sant'AnnaNo ratings yet
- Biomarkers Related To Endometrial Receptivity and ImplantationDocument19 pagesBiomarkers Related To Endometrial Receptivity and ImplantationEndriana Gak Endut EndutNo ratings yet
- Nihms 254232Document24 pagesNihms 254232Gabiich AlvaradoNo ratings yet
- 97 FullDocument9 pages97 FullFarasha RashaNo ratings yet
- Review Adipokines in The Skeleton: Influence On Cartilage Function and Joint Degenerative DiseasesDocument8 pagesReview Adipokines in The Skeleton: Influence On Cartilage Function and Joint Degenerative DiseasesLinda FatrisiaNo ratings yet
- The Hormonal Control of Food IntakeDocument23 pagesThe Hormonal Control of Food IntakeFarin MauliaNo ratings yet
- The Effect of Bisphenol A On Puberty: A Critical Review of The Medical LiteratureDocument20 pagesThe Effect of Bisphenol A On Puberty: A Critical Review of The Medical Literaturethe dayNo ratings yet
- Frontiers in Neuroendocrinology: Tony M. PlantDocument16 pagesFrontiers in Neuroendocrinology: Tony M. PlantRUBY JOJOANo ratings yet
- 92NYAS Robinson&smothermanDocument31 pages92NYAS Robinson&smothermanScott RobinsonNo ratings yet
- Animreprod-15-Supplement 1-1003Document12 pagesAnimreprod-15-Supplement 1-1003Thiago LoloNo ratings yet
- Neuronendocrine Control of The Onset of Puberty 2015 PlantDocument16 pagesNeuronendocrine Control of The Onset of Puberty 2015 PlantCarlos MartínezNo ratings yet
- Leptin: Regulation and Clinical ApplicationsFrom EverandLeptin: Regulation and Clinical ApplicationsSam Dagogo-Jack, MDNo ratings yet
- Puberty. Menopause. Women's Ageing: Manuela RussuDocument41 pagesPuberty. Menopause. Women's Ageing: Manuela RussuAnonymous 0XqZUl06PmNo ratings yet
- NutrientsDocument34 pagesNutrientssilvanaNo ratings yet
- Epigenetics in The Extreme: Prions and The Inheritance of Environmentally Acquired Traits (A Critique)Document2 pagesEpigenetics in The Extreme: Prions and The Inheritance of Environmentally Acquired Traits (A Critique)Daniel Dela CruzNo ratings yet
- 2015 Embryos, DOHaD and David BarkerDocument7 pages2015 Embryos, DOHaD and David BarkerNguyễn Tiến HồngNo ratings yet
- Genetic and Epigenetic Causes of ObesityDocument26 pagesGenetic and Epigenetic Causes of ObesityBudi Haryadi PNo ratings yet
- The Role of Leptin and Ghrelin in The Regulation of Food Intake and Body Weight in Humans: A ReviewDocument14 pagesThe Role of Leptin and Ghrelin in The Regulation of Food Intake and Body Weight in Humans: A ReviewKim Nana UlfaNo ratings yet
- Nutrigenomics in Livestock - Recent AdvancesDocument11 pagesNutrigenomics in Livestock - Recent AdvancesIpung ArmstrongNo ratings yet
- Gut-Microbiome-Obesity-And-Metabolic-DysfunctionDocument8 pagesGut-Microbiome-Obesity-And-Metabolic-DysfunctionSandraosoNo ratings yet
- The Prevention of Aging in MammalsDocument5 pagesThe Prevention of Aging in MammalsJeremy KnippleNo ratings yet
- Scaramuzzi 2008Document8 pagesScaramuzzi 2008Canuto MunozNo ratings yet
- Interaksi Endokrin Dalam Pengaturan Pertumbuhan JaninDocument40 pagesInteraksi Endokrin Dalam Pengaturan Pertumbuhan JaninFeni RiandariNo ratings yet
- Reconocimiento Materno de EmbrionDocument15 pagesReconocimiento Materno de Embrionportillajesus17No ratings yet
- Eje 257Document15 pagesEje 257Sam SoeteNo ratings yet
- P4 Uso Clinico 2015Document11 pagesP4 Uso Clinico 2015Cinthia DarceNo ratings yet
- Reproduction: Role of Angiotensin in Ovarian Follicular Development and Ovulation in Mammals: A Review of Recent AdvancesDocument10 pagesReproduction: Role of Angiotensin in Ovarian Follicular Development and Ovulation in Mammals: A Review of Recent AdvancesWalter Zuta MenaNo ratings yet
- Gynecology and Minimally Invasive Therapy: Case ReportDocument5 pagesGynecology and Minimally Invasive Therapy: Case ReportelenNo ratings yet
- A Review Leptin Structure and Mechanism Actions"Document8 pagesA Review Leptin Structure and Mechanism Actions"elenNo ratings yet
- 03 Bartholin Gland Procedure + Inform ConsentDocument4 pages03 Bartholin Gland Procedure + Inform ConsentelenNo ratings yet
- Thyroid Autoimmunity and Female Infertility: Kris Poppe, Daniel Glinoer, Brigitte VelkeniersDocument16 pagesThyroid Autoimmunity and Female Infertility: Kris Poppe, Daniel Glinoer, Brigitte VelkenierselenNo ratings yet
- Drainage of Bartholin Cyst Abscess ML46071Document4 pagesDrainage of Bartholin Cyst Abscess ML46071elenNo ratings yet
- Thyroid Autoimmunity and Female Infertility: Kris Poppe, Daniel Glinoer, Brigitte VelkeniersDocument16 pagesThyroid Autoimmunity and Female Infertility: Kris Poppe, Daniel Glinoer, Brigitte VelkenierselenNo ratings yet
- Severe Preeclampsia BahanDocument7 pagesSevere Preeclampsia BahanelenNo ratings yet
- Alo AjogDocument1 pageAlo AjogelenNo ratings yet
- Best Practice in Comprehensive Abortion Care RCOGDocument17 pagesBest Practice in Comprehensive Abortion Care RCOGelenNo ratings yet
- 294 Fetal Heart Rate Monitoring. Principles and Interpretation of Cardiotocography LR PDFDocument9 pages294 Fetal Heart Rate Monitoring. Principles and Interpretation of Cardiotocography LR PDFelenNo ratings yet
- Abdul Qudus PatologiDocument37 pagesAbdul Qudus PatologiAgus SetiadiNo ratings yet
- Klinik "MMC": Harga Jual Laboratorium Jln. Budi UtomoDocument1 pageKlinik "MMC": Harga Jual Laboratorium Jln. Budi UtomoNikmah's Dentist LifeNo ratings yet
- Robert Crown Michael Script Puberatate BaietiDocument22 pagesRobert Crown Michael Script Puberatate BaietiElena GheoculescuNo ratings yet
- ARDANADocument7 pagesARDANAArben Anthony Saavedra QuitosNo ratings yet
- Gynecology - Anatomy: Dr. Fidia MumtahanaDocument125 pagesGynecology - Anatomy: Dr. Fidia MumtahanaSonny JhaNo ratings yet
- A Case Study: Region 2 University of La Salette, Inc. Santiago City, IsabelaDocument11 pagesA Case Study: Region 2 University of La Salette, Inc. Santiago City, IsabelaronronNo ratings yet
- Genetic TechnologyDocument5 pagesGenetic TechnologyAnuradha BudhathokiNo ratings yet
- Biology 5090 12 Paper 1 Question Paper Mayjue 2011Document20 pagesBiology 5090 12 Paper 1 Question Paper Mayjue 2011Raja Muda Raja NgahNo ratings yet
- Diasino® One-Step Sandwich Anti-Mullerian Hormone (Amh) AssayDocument4 pagesDiasino® One-Step Sandwich Anti-Mullerian Hormone (Amh) AssayAbdalazeez AlsayedNo ratings yet
- Pcod FnalDocument7 pagesPcod FnalpbalasankarNo ratings yet
- Dystocia Due To Bull Dog Foetal Monster Calf in A Buffaloe PDFDocument1 pageDystocia Due To Bull Dog Foetal Monster Calf in A Buffaloe PDFRavi RaidurgNo ratings yet
- Development of Pituitary Gland From Pices To MammaliansDocument18 pagesDevelopment of Pituitary Gland From Pices To MammaliansTalat naseemNo ratings yet
- Wound DebridementDocument9 pagesWound Debridementannamariaangeles0% (2)
- Cestodes: - Phylum Platyhelminthes (Flatworms)Document23 pagesCestodes: - Phylum Platyhelminthes (Flatworms)ruben6mNo ratings yet
- Igcse Biology Section 3 Lesson 4Document74 pagesIgcse Biology Section 3 Lesson 4sheelaNo ratings yet
- Leopold's Maneuver SCRPTDocument1 pageLeopold's Maneuver SCRPTDarwin QuirimitNo ratings yet
- Van Haute PDFDocument16 pagesVan Haute PDFMads Peter KarlsenNo ratings yet
- Occupational Health Status of Workers Exposed To Female Hormones in A Pharmaceutical PlantDocument6 pagesOccupational Health Status of Workers Exposed To Female Hormones in A Pharmaceutical Plantprom80117No ratings yet
- Skor Bishop, Profil Biofisik Janin, Dan Tanda Kehamilan Post-TermDocument9 pagesSkor Bishop, Profil Biofisik Janin, Dan Tanda Kehamilan Post-TermMuhammad KhoiruddinNo ratings yet
- Family Research Institute - Educational PamphletsDocument61 pagesFamily Research Institute - Educational Pamphletssavoisien88No ratings yet
- Bunt - Male and Female Infertility USAFP 2009-v2 (PPTminimizer)Document35 pagesBunt - Male and Female Infertility USAFP 2009-v2 (PPTminimizer)Jalajarani AridassNo ratings yet
- CHAPTER 6 Animalia (Invertebrate)Document17 pagesCHAPTER 6 Animalia (Invertebrate)mottong100% (1)
- PestsDocument41 pagesPestsFarai FaustosNo ratings yet
- Fertility Control in Soviet Russia, 1920-1936Document268 pagesFertility Control in Soviet Russia, 1920-1936LilianaNo ratings yet
- Quiz 9 Human Reproduction Pregnancy Menstruation GraphsDocument4 pagesQuiz 9 Human Reproduction Pregnancy Menstruation GraphsCamille FrancoNo ratings yet
- Kualitas Hidup MenopauseDocument9 pagesKualitas Hidup MenopauseRoviqi Bozgank AzemNo ratings yet
- Sexually Transmitted Diseases (STDS) Tahir Kundki 6Document22 pagesSexually Transmitted Diseases (STDS) Tahir Kundki 6shahrukhziaNo ratings yet
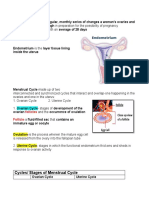
- Cycles/ Stages of Menstrual Cycle: Follicles FollicleDocument6 pagesCycles/ Stages of Menstrual Cycle: Follicles FollicleAaliyah CarlobosNo ratings yet
- Epithelial Ovarian CancerDocument14 pagesEpithelial Ovarian Cancernautilus81No ratings yet