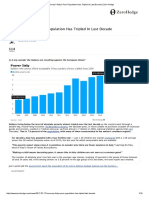
Download as pdf or txt
You might also like
- S4C01E - SAP S - 4HANA Cloud On-Boarding Fundamentals - SAP TrainingDocument4 pagesS4C01E - SAP S - 4HANA Cloud On-Boarding Fundamentals - SAP Trainingfiestamix13% (8)
- SCM600 - Sales Order Management - SAP TrainingDocument7 pagesSCM600 - Sales Order Management - SAP TrainingfiestamixNo ratings yet
- MDG100 - SAP Master Data Governance - SAP TrainingDocument7 pagesMDG100 - SAP Master Data Governance - SAP Trainingfiestamix0% (3)
- The New Mind Readers: What Neuroimaging Can and Cannot Reveal about Our ThoughtsFrom EverandThe New Mind Readers: What Neuroimaging Can and Cannot Reveal about Our ThoughtsNo ratings yet
- SCM605 - Sales Processing in SAP ERP - SAP TrainingDocument7 pagesSCM605 - Sales Processing in SAP ERP - SAP Trainingfiestamix33% (3)
- Raven Prabhakaran1997 PDFDocument21 pagesRaven Prabhakaran1997 PDFargiaescuNo ratings yet
- Aphasia RehabDocument72 pagesAphasia RehabkicsirekaNo ratings yet
- Neural Substrates of Fluid Reasoning: An fMRI Study of Neocortical Activation During Performance of The Raven's Progressive Matrices TestDocument21 pagesNeural Substrates of Fluid Reasoning: An fMRI Study of Neocortical Activation During Performance of The Raven's Progressive Matrices TestAviraj MahadikNo ratings yet
- Berman Et Al., 2006 Studying Mind and Brain With fMRIDocument4 pagesBerman Et Al., 2006 Studying Mind and Brain With fMRIMarkNo ratings yet
- NIH Public Access: Future Trends in Neuroimaging: Neural Processes As Expressed Within Real-Life ContextsDocument14 pagesNIH Public Access: Future Trends in Neuroimaging: Neural Processes As Expressed Within Real-Life ContextslitrevnmNo ratings yet
- Do We Need To Study The Brain To Understand The MindDocument3 pagesDo We Need To Study The Brain To Understand The MindMARCO ANTONIO OLIVARES HURTADONo ratings yet
- Theoretical, Statistical, and Practical Perspectives On Pattern-Based Classification Approaches To The Analysis of Functional Neuroimaging DataDocument19 pagesTheoretical, Statistical, and Practical Perspectives On Pattern-Based Classification Approaches To The Analysis of Functional Neuroimaging DataNoScriptNo ratings yet
- 1 Functional Neuroimaging of Executive Functions Todd S. Braver and Hannes Ruge Department ... (PDFDrive)Document93 pages1 Functional Neuroimaging of Executive Functions Todd S. Braver and Hannes Ruge Department ... (PDFDrive)Amornchai WongworrakunNo ratings yet
- Contributions of Episodic Retrieval and Mentalizing To Autobiographical ThoughtDocument12 pagesContributions of Episodic Retrieval and Mentalizing To Autobiographical ThoughtFrancisco Ahumada MéndezNo ratings yet
- ConsciousnessDocument4 pagesConsciousnessarhodes777No ratings yet
- The Multiple Functions of Sensory-Motor (2005)Document3 pagesThe Multiple Functions of Sensory-Motor (2005)ANTONIO REYES MEDINANo ratings yet
- Soc Cogn Affect Neurosci 2016 Greven 641 51Document11 pagesSoc Cogn Affect Neurosci 2016 Greven 641 51tefsNo ratings yet
- MVPA Personality - PreprintDocument14 pagesMVPA Personality - PreprintGeo MeNo ratings yet
- The Brain's Default Network: Anatomy, Function, and Relevance To DiseaseDocument38 pagesThe Brain's Default Network: Anatomy, Function, and Relevance To DiseaseMarkoff ChaneyNo ratings yet
- Facial PerceptionDocument7 pagesFacial PerceptionGECJNo ratings yet
- Vinod Goel (2007) - Anatomy of Deductive ReasoningDocument7 pagesVinod Goel (2007) - Anatomy of Deductive ReasoningRodrigo CórdovaNo ratings yet
- Plasticité Nrpsy CogDocument14 pagesPlasticité Nrpsy CogMalak GabsiNo ratings yet
- Depisapia AISC2010Document5 pagesDepisapia AISC2010Nicola De PisapiaNo ratings yet
- Image Search AlgorithmDocument28 pagesImage Search AlgorithmbinzbinzNo ratings yet
- Fnhum 07 00692Document6 pagesFnhum 07 00692bronson.harryNo ratings yet
- Reitan 1994Document38 pagesReitan 1994Sergio Machado NeurocientistaNo ratings yet
- Nancy - Face AreaDocument20 pagesNancy - Face AreaOchopinochoNo ratings yet
- Nihms 474787Document7 pagesNihms 474787ClaraCalvoTorresNo ratings yet
- J Neuroimage 2009 12 098Document8 pagesJ Neuroimage 2009 12 098hanbing yinNo ratings yet
- Executive Functions As Virtual Machines: A New Metaphor For A Neuropsychological Classic ConceptDocument11 pagesExecutive Functions As Virtual Machines: A New Metaphor For A Neuropsychological Classic ConceptsiscupNo ratings yet
- En Lo AmbientalDocument11 pagesEn Lo AmbientalYocelin Espiritu ReynosoNo ratings yet
- Spunt (2011) JOCNDocument12 pagesSpunt (2011) JOCNBUBUSNo ratings yet
- What Can Be Learned About Normal Cognitive Function Using Haemodynamic Imaging Techniques?Document6 pagesWhat Can Be Learned About Normal Cognitive Function Using Haemodynamic Imaging Techniques?Stephen LissendenNo ratings yet
- Superfluous Neuroscience Information Makes Explanations of Psychological Phenomena More AppealingDocument19 pagesSuperfluous Neuroscience Information Makes Explanations of Psychological Phenomena More AppealingBrianNo ratings yet
- Self-Reported Empathy and Neural Activity During Action Imitation andDocument9 pagesSelf-Reported Empathy and Neural Activity During Action Imitation andPheeraphol ThapprasithiNo ratings yet
- Mind The Brain Mind The Brain Mind The Brain Mind The BrainDocument4 pagesMind The Brain Mind The Brain Mind The Brain Mind The BrainMariø YŋgveøhliŋNo ratings yet
- Vivian Wu 260891725 Paper 6Document3 pagesVivian Wu 260891725 Paper 6Vivian WuNo ratings yet
- Affective Mapping An Activation Likelihood Estimation (ALE) Meta-AnalysisDocument12 pagesAffective Mapping An Activation Likelihood Estimation (ALE) Meta-Analysisselina hahnNo ratings yet
- Neural Correlates of The Object-Recall Process in Semantic MemoryDocument12 pagesNeural Correlates of The Object-Recall Process in Semantic MemoryMilenaNo ratings yet
- A Default Mode of Brain Function - A Brief History of An Evolving IdeaDocument8 pagesA Default Mode of Brain Function - A Brief History of An Evolving IdeaEnzimoshNo ratings yet
- An Introduction To Anatomical ROI-based fMRI Classification AnalysisDocument12 pagesAn Introduction To Anatomical ROI-based fMRI Classification AnalysisFridRachmanNo ratings yet
- What Is Self SpecificDocument31 pagesWhat Is Self SpecificchandusgNo ratings yet
- NIH Public Access: Author ManuscriptDocument18 pagesNIH Public Access: Author ManuscriptYeremias EdwinNo ratings yet
- 114-2005 General Specific NeuroImageDocument6 pages114-2005 General Specific NeuroImageEdward Ventura BarrientosNo ratings yet
- Jurnal 2Document10 pagesJurnal 2akreditasimedicalpegadaianNo ratings yet
- Tunik Et Al. 2007Document10 pagesTunik Et Al. 2007Miguel Antonio Barretto GarcíaNo ratings yet
- Abernathy Dylan Context Memo Cover Letter 1 KF 3Document3 pagesAbernathy Dylan Context Memo Cover Letter 1 KF 3api-710551120No ratings yet
- Understanding SelfDocument9 pagesUnderstanding SelfNaja ReyesNo ratings yet
- Burgess 08 SpatialDocument21 pagesBurgess 08 Spatialsarabiba1971No ratings yet
- Cognitive NeuroscienceDocument4 pagesCognitive NeuroscienceAkhwand SaulatNo ratings yet
- Hemispheric Reasoning StrategiesDocument10 pagesHemispheric Reasoning Strategiesrodrigo_rrNo ratings yet
- Editorial: J.Neurol Sci (Turk)Document4 pagesEditorial: J.Neurol Sci (Turk)Nezih OktarNo ratings yet
- Current Topics in Cognitive Psychology, 2010Document15 pagesCurrent Topics in Cognitive Psychology, 2010Hanne Melgård WatkinsNo ratings yet
- Pre Mid Assignment 1 Sania Saddique (Book)Document7 pagesPre Mid Assignment 1 Sania Saddique (Book)Sania SaddiqueNo ratings yet
- VanOverwalle2020 Article ConsensusPaperCerebellumAndSocDocument36 pagesVanOverwalle2020 Article ConsensusPaperCerebellumAndSocTSGNo ratings yet
- Decision Making Brain Rorie 2005Document3 pagesDecision Making Brain Rorie 2005neurocastedNo ratings yet
- Cognitive Synthesis TestDocument33 pagesCognitive Synthesis TestvedroconmioNo ratings yet
- Self-Esteem Modulates Dorsal Medial Prefrontal Cortical Response To Self-Positivity Bias in Implicit Self-Relevant ProcessingDocument5 pagesSelf-Esteem Modulates Dorsal Medial Prefrontal Cortical Response To Self-Positivity Bias in Implicit Self-Relevant ProcessingJoeNo ratings yet
- HHS Public Access: Neural Architecture Underlying Classification of Face Perception ParadigmsDocument26 pagesHHS Public Access: Neural Architecture Underlying Classification of Face Perception Paradigmscecilia martinezNo ratings yet
- fMRI Analysis of Situational and Behavior Description Interview QuestionsDocument12 pagesfMRI Analysis of Situational and Behavior Description Interview QuestionsEszter SzerencsiNo ratings yet
- 2008 Investigating Paranormal Phenomena Functional Brain Imaging of TelepathyDocument6 pages2008 Investigating Paranormal Phenomena Functional Brain Imaging of TelepathymarciascognamilloNo ratings yet
- 2004 Towards Studies of The Social BrainDocument5 pages2004 Towards Studies of The Social BrainAl WilsonNo ratings yet
- Neural Reuse Anderson 2010Document69 pagesNeural Reuse Anderson 2010Katie BanksNo ratings yet
- The Brain has a Mind of its Own: Attachment, Neurobiology, and the New Science of PsychotherapyFrom EverandThe Brain has a Mind of its Own: Attachment, Neurobiology, and the New Science of PsychotherapyNo ratings yet
- Stock Markets Still in The Doldrums - BBC NewsDocument17 pagesStock Markets Still in The Doldrums - BBC NewsfiestamixNo ratings yet
- What'Ll Happen To US Commercial Real Estate As Chinese Money Dries Up - Zero HedgeDocument6 pagesWhat'Ll Happen To US Commercial Real Estate As Chinese Money Dries Up - Zero HedgefiestamixNo ratings yet
- No-Deal Brexit 'To Leave Shelves Empty' Warn Retailers - BBC NewsDocument14 pagesNo-Deal Brexit 'To Leave Shelves Empty' Warn Retailers - BBC NewsfiestamixNo ratings yet
- ? - 40 @abc BB ? - 40 @ibc BBDocument4 pages? - 40 @abc BB ? - 40 @ibc BBfiestamixNo ratings yet
- SapDocument4 pagesSapfiestamixNo ratings yet
- Unit 1: Introduction To Webdynpro ABAPDocument43 pagesUnit 1: Introduction To Webdynpro ABAPfiestamixNo ratings yet
- Which Is A Good Book For SAP SD - QuoraDocument2 pagesWhich Is A Good Book For SAP SD - QuorafiestamixNo ratings yet
- The Mistake That Led To A 1.2bn Business - BBC NewsDocument14 pagesThe Mistake That Led To A 1.2bn Business - BBC NewsfiestamixNo ratings yet
- SAP SD Certification Exam Syllabus - ERPPrepDocument4 pagesSAP SD Certification Exam Syllabus - ERPPrepfiestamix0% (1)
- Curriculum - Application (Solution) Consultant SAP SCM - SAP ERP - Order Fulfillment (Sales Order Management)Document3 pagesCurriculum - Application (Solution) Consultant SAP SCM - SAP ERP - Order Fulfillment (Sales Order Management)fiestamixNo ratings yet
- Recovery - Italy's Poor Population Has Tripled in Last Decade - Zero HedgeDocument19 pagesRecovery - Italy's Poor Population Has Tripled in Last Decade - Zero HedgefiestamixNo ratings yet
- SCM610 - Delivery Processing in SAP ERP - SAP TrainingDocument7 pagesSCM610 - Delivery Processing in SAP ERP - SAP TrainingfiestamixNo ratings yet
- American Students Love Socialism (Just Don't Ask Them What It Is) - Zero HedgeDocument22 pagesAmerican Students Love Socialism (Just Don't Ask Them What It Is) - Zero HedgefiestamixNo ratings yet
- SCM210 - Core Interface (CIF) - SAP TrainingDocument5 pagesSCM210 - Core Interface (CIF) - SAP TrainingfiestamixNo ratings yet
- SCM610 - Delivery Processing in SAP ERP - SAP TrainingDocument7 pagesSCM610 - Delivery Processing in SAP ERP - SAP TrainingfiestamixNo ratings yet
- TEWM10 - Extended Warehouse Management I - SAP TrainingDocument4 pagesTEWM10 - Extended Warehouse Management I - SAP TrainingfiestamixNo ratings yet
- S4F00E - Overview of Financials in SAP S - 4HANA - SAP Training PDFDocument3 pagesS4F00E - Overview of Financials in SAP S - 4HANA - SAP Training PDFfiestamix0% (1)
- S4F00E - Overview of Financials in SAP S - 4HANA - SAP TrainingDocument3 pagesS4F00E - Overview of Financials in SAP S - 4HANA - SAP TrainingfiestamixNo ratings yet
- Only in San Francisco - Couples Making $138k A Year Now Qualify For Subsidized - Affordable Housing - Zero HedgeDocument13 pagesOnly in San Francisco - Couples Making $138k A Year Now Qualify For Subsidized - Affordable Housing - Zero HedgefiestamixNo ratings yet
- SAP Easy Access - SAP Training and CertificationDocument16 pagesSAP Easy Access - SAP Training and Certificationfiestamix100% (1)
- SAP FI Certification (Financial Accounting) - SAP CertificationDocument5 pagesSAP FI Certification (Financial Accounting) - SAP CertificationfiestamixNo ratings yet
- SAP Super User - SAP Training and CertificationDocument5 pagesSAP Super User - SAP Training and CertificationfiestamixNo ratings yet
- Connectivism and Connective KnowledgeDocument616 pagesConnectivism and Connective KnowledgeleeyikshengNo ratings yet
- Appraisal PointsDocument2 pagesAppraisal PointsPiyasha NathNo ratings yet
- CEL2102 Project 1 - DescriptionDocument6 pagesCEL2102 Project 1 - DescriptionAbdul Qhouyyum AiedilNo ratings yet
- The Rise of Digital Intelligence Challenges For Public Relations Education and PracticesDocument18 pagesThe Rise of Digital Intelligence Challenges For Public Relations Education and PracticesTiana AlexeNo ratings yet
- Coaching in Human ResourceDocument21 pagesCoaching in Human Resourcesya_ezNo ratings yet
- QuestionsDocument20 pagesQuestionsapi-295210059No ratings yet
- The Importance of Discomfort On NoFapDocument4 pagesThe Importance of Discomfort On NoFapVAXAEVNo ratings yet
- Senior High School: Organization & Management Quarter IIDocument18 pagesSenior High School: Organization & Management Quarter IICipher Amoz100% (1)
- Final OUTLINE Art&CreativityDocument11 pagesFinal OUTLINE Art&CreativityJohanna GutierrezNo ratings yet
- Mult Intelligence QuizDocument2 pagesMult Intelligence Quizapi-299481084No ratings yet
- Free Sample of An Action Research ProposalDocument18 pagesFree Sample of An Action Research Proposalreginald_adia_1No ratings yet
- Theory Part 4Document4 pagesTheory Part 4Abad ChristineNo ratings yet
- ETHICSDocument7 pagesETHICSErlan Grace HeceraNo ratings yet
- Creative As A Prerequisite To InnovationDocument2 pagesCreative As A Prerequisite To InnovationSaad Rehman100% (2)
- WORKING THESIS (Teavher's Effectiveness in Teaching The English Language As Perceived by The Grade 9 Students of Quezonian Educational College Inc., Atimonan, Quezon S.Y. 2017-2018Document54 pagesWORKING THESIS (Teavher's Effectiveness in Teaching The English Language As Perceived by The Grade 9 Students of Quezonian Educational College Inc., Atimonan, Quezon S.Y. 2017-2018Christofer De Los ReyesNo ratings yet
- Cls 6Document5 pagesCls 6popalina24No ratings yet
- Relationship Between Reading and Writing Performance (Rosita Aminullah) PP 179-182Document4 pagesRelationship Between Reading and Writing Performance (Rosita Aminullah) PP 179-182upenapahangNo ratings yet
- Hall - S The Work of RepresentationDocument30 pagesHall - S The Work of RepresentationmarleneprestesNo ratings yet
- Marketing Keller's Brand Equity ModelDocument3 pagesMarketing Keller's Brand Equity ModelSana Vancy100% (1)
- Mind HackDocument6 pagesMind Hackapi-374080275% (4)
- Module 5 - The Emotional SelfDocument5 pagesModule 5 - The Emotional SelfMonasque PamelaNo ratings yet
- Running Head: Interviewing Types and TechniquesDocument6 pagesRunning Head: Interviewing Types and Techniquesklm klmNo ratings yet
- Week 1 Undefined TermsDocument8 pagesWeek 1 Undefined TermsJo-Amver Valera ManzanoNo ratings yet
- Tweed 2020 Faith in HumanityDocument13 pagesTweed 2020 Faith in Humanitymuiescr1bdNo ratings yet
- By Liam Lindsay: Cover Photo by Andrea RothDocument32 pagesBy Liam Lindsay: Cover Photo by Andrea RothSandraNo ratings yet
- Critical ThikingDocument17 pagesCritical ThikingLen AungNo ratings yet
- Ethic in Public SectorDocument46 pagesEthic in Public SectorCOT Management Training Insitute100% (2)
- Writing Cases: Melodena Stephens BalakrishnanDocument6 pagesWriting Cases: Melodena Stephens BalakrishnanErika L.No ratings yet
- What Is Divided Attention?Document50 pagesWhat Is Divided Attention?LoraNo ratings yet
- Problem Solving StrategiesDocument3 pagesProblem Solving StrategiesJA M ESNo ratings yet