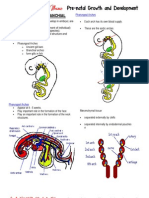
Download as docx, pdf, or txt
You might also like
- Yousaf Ali (Auth.) - Self Assessment Questions in Rheumatology-Humana Press (2009)Document147 pagesYousaf Ali (Auth.) - Self Assessment Questions in Rheumatology-Humana Press (2009)Amr AmalNo ratings yet
- Mens Fitness UK - March 2016Document180 pagesMens Fitness UK - March 2016Raphael Doukkali100% (4)
- Anatomy of Cardiovascular SystemDocument23 pagesAnatomy of Cardiovascular SystemElvira Wisakhadewi100% (1)
- CHAPTER 3 GcsDocument14 pagesCHAPTER 3 GcsNurse NotesNo ratings yet
- Istology: Lymphatic SystemDocument48 pagesIstology: Lymphatic Systemninja-2001No ratings yet
- Cardiac Cycle - DR Rakesh JainDocument97 pagesCardiac Cycle - DR Rakesh JainEmmieNo ratings yet
- Development of Branchial ArchesDocument4 pagesDevelopment of Branchial ArchesFidz Lianko100% (1)
- Progenitor Heart Cells Lie in The Epiblast, Immediately Adjacent To TheDocument15 pagesProgenitor Heart Cells Lie in The Epiblast, Immediately Adjacent To TheJehangir AllamNo ratings yet
- Anatomical Terms: by WWW - Ipostforyou.infoDocument38 pagesAnatomical Terms: by WWW - Ipostforyou.infoShahid Maqbool100% (1)
- Human Anatomy,: First Edition Mckinley & O'LoughlinDocument40 pagesHuman Anatomy,: First Edition Mckinley & O'LoughlinAgung PurmanaNo ratings yet
- Large IntestineDocument50 pagesLarge Intestinemex Gbrekorkos100% (1)
- Anatomy of Larynx Lecture 7Document21 pagesAnatomy of Larynx Lecture 7sallykamareddine100% (1)
- Lymphatic System and ImmunityDocument5 pagesLymphatic System and ImmunityRheeanne AmilasanNo ratings yet
- Gross Anatomy of Posterior Compartment of The ThighDocument20 pagesGross Anatomy of Posterior Compartment of The ThighOnah ArinzeNo ratings yet
- The Circulatory SystemDocument8 pagesThe Circulatory SystembysshelarbyNo ratings yet
- 1.anatomy of The PleuraDocument17 pages1.anatomy of The PleuraMangayakkarasi Telukanam0% (1)
- Structure and Functions of The Major Types of Blood VesselsDocument6 pagesStructure and Functions of The Major Types of Blood VesselsCharls DeimoyNo ratings yet
- Chapter 9 - Muscular System: Three Types of Muscle TissuesDocument73 pagesChapter 9 - Muscular System: Three Types of Muscle TissuesRafie HusniNo ratings yet
- Digestive System - MefloresDocument117 pagesDigestive System - MefloresJohnMichaelDominguezNo ratings yet
- Composition and Function of Blood ComponentsDocument17 pagesComposition and Function of Blood ComponentsPrakash Kumar Nayak100% (1)
- The Pelvic Girdle: By/Dr - Abdisamad Omar AliDocument30 pagesThe Pelvic Girdle: By/Dr - Abdisamad Omar AliSAKARIYE MAXAMEDNo ratings yet
- Cardiovascular System: The Heart: Chapter 19 - Lecture NotesDocument73 pagesCardiovascular System: The Heart: Chapter 19 - Lecture Noteshersheys72002No ratings yet
- Cardiovascular System: K. Hariharan Iv Eee - 'B'Document33 pagesCardiovascular System: K. Hariharan Iv Eee - 'B'Hari Haran100% (1)
- Circulatory SystemDocument30 pagesCirculatory SystemIra Lavigne De VillaNo ratings yet
- Physiology of The Cardiovascular System-CVSDocument56 pagesPhysiology of The Cardiovascular System-CVSAmanuel MaruNo ratings yet
- MediastinumDocument24 pagesMediastinumapi-249972919100% (1)
- Tissues 4Document65 pagesTissues 4marianisie448100% (3)
- Regulation RespirationDocument34 pagesRegulation Respirationkiedd_04100% (2)
- Lect. 11 Respiratory SystemDocument43 pagesLect. 11 Respiratory Systemflex gyNo ratings yet
- Gross Anatomy of LungsDocument8 pagesGross Anatomy of LungsAriguna WijayaNo ratings yet
- Chapter 19 The Circulatory System - The HeartDocument14 pagesChapter 19 The Circulatory System - The HeartGary Bowen100% (1)
- The Cardiovascular System: Group 1Document92 pagesThe Cardiovascular System: Group 1Jefferson AdralesNo ratings yet
- The Thorax Part Ii - The Thoracic Cavity: Juan Guido G. Joyo, PTRP Juvi G. Alicabo, PTRP, CCP, CTMBP, CTTTPDocument98 pagesThe Thorax Part Ii - The Thoracic Cavity: Juan Guido G. Joyo, PTRP Juvi G. Alicabo, PTRP, CCP, CTMBP, CTTTPFerjie Angelica DalandaoNo ratings yet
- Digestive System Power PointDocument17 pagesDigestive System Power PointAlyssa Mae DapadapNo ratings yet
- Embryo Lab Exercise 1Document7 pagesEmbryo Lab Exercise 1Karmina Santos100% (1)
- Angiology NoteDocument13 pagesAngiology NoteSantosh Bhandari100% (2)
- The Neck PDFDocument67 pagesThe Neck PDFAhmad Akram NatshehNo ratings yet
- What Is Electrosurgery?Document4 pagesWhat Is Electrosurgery?RiyaNo ratings yet
- Histology of Tonsil and Splenic Circulation by Dr. RoomiDocument21 pagesHistology of Tonsil and Splenic Circulation by Dr. RoomiMudassar RoomiNo ratings yet
- BIOLAB FROG Organ HistologyDocument7 pagesBIOLAB FROG Organ HistologyCloie Anne Rabinetas100% (1)
- Circulatory System - Part 2 4-8-14 For BBDocument23 pagesCirculatory System - Part 2 4-8-14 For BBroman7dbNo ratings yet
- 16 Sense OrgansDocument79 pages16 Sense OrgansvanderphysNo ratings yet
- Anterior & Medial Compartment of ThighDocument25 pagesAnterior & Medial Compartment of ThighnasibdinNo ratings yet
- 16/09/2015 1 Mujtaba AshrafDocument44 pages16/09/2015 1 Mujtaba Ashrafதீரன் சக்திவேல்No ratings yet
- Cell Cycle and Cell ControlDocument18 pagesCell Cycle and Cell ControlSTANLY MORALESNo ratings yet
- Biochemistry of Blood Elements: The Figure Is Found at (March 2007)Document37 pagesBiochemistry of Blood Elements: The Figure Is Found at (March 2007)Sadam_fasterNo ratings yet
- Lecture 6 Cardiovascular: Vascular System - The HemodynamicsDocument70 pagesLecture 6 Cardiovascular: Vascular System - The HemodynamicsAndreea ŞtefănescuNo ratings yet
- MHC I Proteins, Which Present Antigens To Cytotoxic T Cells, (2) MHC II Proteins, Which Present Antigens T Helper CellsDocument25 pagesMHC I Proteins, Which Present Antigens To Cytotoxic T Cells, (2) MHC II Proteins, Which Present Antigens T Helper CellsMudassar Roomi100% (1)
- The Gut Tube and Body CavitiesDocument31 pagesThe Gut Tube and Body CavitiesGeoffreyNo ratings yet
- Lect 2 Membrane TransportDocument29 pagesLect 2 Membrane Transportramadan100% (1)
- 40 Life Cycle HerdmaniaDocument7 pages40 Life Cycle HerdmaniaAyushiNo ratings yet
- Characteristics of Histology SlidesDocument4 pagesCharacteristics of Histology SlidesLaiq Khan100% (1)
- Clinical Anatomy of ThoraxDocument4 pagesClinical Anatomy of ThoraxCeo MedCom100% (1)
- Heart SoundsDocument23 pagesHeart SoundsK.R.RaguramNo ratings yet
- Lecture Notes: A Powerpoint PresentationDocument105 pagesLecture Notes: A Powerpoint PresentationDutchsMoin Mohammad100% (1)
- Anatomy - Skull FeaturesDocument6 pagesAnatomy - Skull FeaturesAkhil BansalNo ratings yet
- Respiratory Physiology: Dr. Okon MRCSDocument15 pagesRespiratory Physiology: Dr. Okon MRCSEdwin OkonNo ratings yet
- Describe The Factors Affecting Cardiac OutputDocument6 pagesDescribe The Factors Affecting Cardiac OutputSis SukarnoNo ratings yet
- Physiology L6Document6 pagesPhysiology L6Anonymous elq7jZiSNo ratings yet
- Oral RevDocument6 pagesOral RevjjolaguerNo ratings yet
- Management of Clients With Problems of The Cardiovascular SystemDocument7 pagesManagement of Clients With Problems of The Cardiovascular Systemjerry smatt dumtoNo ratings yet
- HUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemDocument12 pagesHUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemZaira MangalimanNo ratings yet
- Mumps Presenting With Unilateral, Synchronous Parotid and Submandibular Gland SwellingDocument4 pagesMumps Presenting With Unilateral, Synchronous Parotid and Submandibular Gland SwellingSamantha Dela CernaNo ratings yet
- ACOG Practice Bulletin ObesityDocument2 pagesACOG Practice Bulletin Obesitynoella deckbarNo ratings yet
- Liver Biopsy: Mary Raina Angeli Fujiyoshi, MDDocument10 pagesLiver Biopsy: Mary Raina Angeli Fujiyoshi, MDRaina FujiyoshiNo ratings yet
- Pulmonary Metastasis 2Document7 pagesPulmonary Metastasis 2raisamentariNo ratings yet
- Malnutrition-Related Diabetes Mellitus in Africa: PrevalenceDocument7 pagesMalnutrition-Related Diabetes Mellitus in Africa: PrevalenceAbdi KebedeNo ratings yet
- English 74 TH Module 8Document12 pagesEnglish 74 TH Module 8Hellen DeaNo ratings yet
- Cebu Doctors' University - College of Medicine Preventive and Social MedicineDocument14 pagesCebu Doctors' University - College of Medicine Preventive and Social MedicinefilchibuffNo ratings yet
- 1,001 Ways To Use Essential Oils - Including 61 Essential Oils - NodrmDocument211 pages1,001 Ways To Use Essential Oils - Including 61 Essential Oils - NodrmFernanda Rodriguez100% (2)
- Jurnal Internasional Kromatografi KolomDocument6 pagesJurnal Internasional Kromatografi KolomRiza FaelasshofaNo ratings yet
- Role of Hematological ParameterDocument7 pagesRole of Hematological ParameterChanif MuyasarohNo ratings yet
- Vaginosis BakterialisDocument9 pagesVaginosis BakterialisIntan PermataNo ratings yet
- Connective TissueDocument2 pagesConnective TissueMarie PetalcorinNo ratings yet
- Regional DialectDocument8 pagesRegional DialectDiana LuncasuNo ratings yet
- Unusual Presentation and Complication of Caustic Ingestion. Case ReportDocument4 pagesUnusual Presentation and Complication of Caustic Ingestion. Case ReportRay MaudyNo ratings yet
- Pilonidal Disease Practice Points: An UpdateDocument3 pagesPilonidal Disease Practice Points: An UpdateNataraj ThambiNo ratings yet
- Home Remedies For HypothyroidismDocument2 pagesHome Remedies For HypothyroidismRatnaPrasadNalamNo ratings yet
- (Download PDF) The Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Full Chapter PDFDocument69 pages(Download PDF) The Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Full Chapter PDFminuredinci100% (7)
- The Oral PresentationDocument6 pagesThe Oral PresentationCarlos MellaNo ratings yet
- List of Autoimmune DiseasesDocument11 pagesList of Autoimmune DiseasesDuwa AhmadNo ratings yet
- Med Surg Bullets 1Document21 pagesMed Surg Bullets 1Jor GarciaNo ratings yet
- Verb Tenses 1 Eval 3º EsoDocument4 pagesVerb Tenses 1 Eval 3º EsoEdurne De Vicente PereiraNo ratings yet
- Common Clinical Manifestations of Gastrointestinal Disease: A. Abdominal PainDocument18 pagesCommon Clinical Manifestations of Gastrointestinal Disease: A. Abdominal PainFranciscoJ.ReynaSepúlvedaNo ratings yet
- Designing A Pilates Program For A Client With Shoulder Impingement SyndromeDocument15 pagesDesigning A Pilates Program For A Client With Shoulder Impingement Syndromebrunoduartefr100% (1)
- Creative Brief For DiabetesDocument6 pagesCreative Brief For DiabetesRahulNo ratings yet
- Health Warning! John Butcher, Surrey Council Councillor Wants To Deny NHS Treatment To His Constituents. Do Not Re-Elect!Document3 pagesHealth Warning! John Butcher, Surrey Council Councillor Wants To Deny NHS Treatment To His Constituents. Do Not Re-Elect!zacharia1206No ratings yet
- Celiac Disease EssayDocument6 pagesCeliac Disease Essaysierra meredithNo ratings yet
- Metabolic Aspects of Chronic Liver DiseaseDocument390 pagesMetabolic Aspects of Chronic Liver DiseaseAndre SamsungNo ratings yet
- P ('t':'3', 'I':'668096276') D '' Var B Location Settimeout (Function ( If (Typeof Window - Iframe 'Undefined') ( B.href B.href ) ), 15000)Document31 pagesP ('t':'3', 'I':'668096276') D '' Var B Location Settimeout (Function ( If (Typeof Window - Iframe 'Undefined') ( B.href B.href ) ), 15000)LipatOla123No ratings yet