
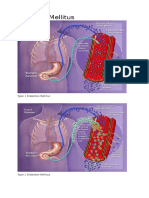
Theoretical Biology and Medical Modelling: A Discrete Single Delay Model For The Intra-Venous Glucose Tolerance Test
Theoretical Biology and Medical Modelling: A Discrete Single Delay Model For The Intra-Venous Glucose Tolerance Test
You might also like
- SJH Newcastle Diet Booklet 2012 Revised 2015Document6 pagesSJH Newcastle Diet Booklet 2012 Revised 2015TexasPedi100% (1)
- tmp22D3 TMPDocument9 pagestmp22D3 TMPFrontiersNo ratings yet
- RM ProtocolDocument4 pagesRM Protocolcifha diasNo ratings yet
- Thorough Testing of Predictive Functional Control On A Well Validated in Silico Model of Type I Diabetic PatientsDocument12 pagesThorough Testing of Predictive Functional Control On A Well Validated in Silico Model of Type I Diabetic PatientsgercampeNo ratings yet
- Manuscript 24092017 UcldDocument46 pagesManuscript 24092017 UcldPrima Hari NastitiNo ratings yet
- Divya 13Document20 pagesDivya 13Divya NadarNo ratings yet
- Song 2014Document11 pagesSong 2014Ayssa WitjaksonoNo ratings yet
- Journal Pone 0248280 PDFDocument17 pagesJournal Pone 0248280 PDFHafis KafitNo ratings yet
- Measurement of Glucose Homeostasis in VivoDocument10 pagesMeasurement of Glucose Homeostasis in Vivo19400668No ratings yet
- Clinical Study: Obesity and Cytokines in Childhood-Onset Systemic Lupus ErythematosusDocument6 pagesClinical Study: Obesity and Cytokines in Childhood-Onset Systemic Lupus ErythematosusIndra KusumayadiNo ratings yet
- Article AmalDocument16 pagesArticle Amalsaliha mharchiNo ratings yet
- InositolDocument14 pagesInositolEdna De Azevedo GabrielNo ratings yet
- Estimating Insulin Sensitivity and Beta-Cell Function From The Oral Glucose Tolerance Test (Ha Et Al., 2024)Document47 pagesEstimating Insulin Sensitivity and Beta-Cell Function From The Oral Glucose Tolerance Test (Ha Et Al., 2024)Marcelo TrNo ratings yet
- Association of Coagulation Profile With Microvascular Complications and Glycemic Control in Type 2 Diabetes Mellitus - A Study ADocument1 pageAssociation of Coagulation Profile With Microvascular Complications and Glycemic Control in Type 2 Diabetes Mellitus - A Study Aseneh17198No ratings yet
- Continuous Glucose Monitoring Devices For Patients With Diabeles Mellitus On InsulinDocument25 pagesContinuous Glucose Monitoring Devices For Patients With Diabeles Mellitus On InsulinCalifornia Technology Assessment ForumNo ratings yet
- 11 Effect Probiotics On Glucose MetabolismDocument7 pages11 Effect Probiotics On Glucose Metabolismbangd1f4nNo ratings yet
- Diabetes Research and Clinical Practice Volume Issue 2018Document37 pagesDiabetes Research and Clinical Practice Volume Issue 2018yayan_adiutama98No ratings yet
- First-Trimester Prediction of Gestational Diabetes Mellitus: Examining The Potential of Combining Maternal Characteristics and Laboratory MeasuresDocument6 pagesFirst-Trimester Prediction of Gestational Diabetes Mellitus: Examining The Potential of Combining Maternal Characteristics and Laboratory MeasuresLucky PratamaNo ratings yet
- Glycemic 2010 PDFDocument9 pagesGlycemic 2010 PDFSiva SankarNo ratings yet
- Glutamina IVDocument11 pagesGlutamina IVBLADIMIR ALEJANDRO GIL VALENCIANo ratings yet
- Human Insulin Vs Insulin AnalogDocument7 pagesHuman Insulin Vs Insulin AnalogWita Ferani Kartika100% (1)
- Tight Glycemic Control in Critically Ill PediatricDocument11 pagesTight Glycemic Control in Critically Ill Pediatricbella friscaamaliaNo ratings yet
- Gene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GeneDocument7 pagesGene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GenesaifudinNo ratings yet
- Associations Between The MetAssociations Between The Metabolic Syndrome Abolic Syndrome andDocument9 pagesAssociations Between The MetAssociations Between The Metabolic Syndrome Abolic Syndrome andEsteban Avila IraguenNo ratings yet
- Time-Delay Model-Based Control of The Glucose-Insulin System, by Means of A State ObserverDocument16 pagesTime-Delay Model-Based Control of The Glucose-Insulin System, by Means of A State ObserverChanachai PuckNo ratings yet
- Palumbo 13Document13 pagesPalumbo 13Margarete Ap SantosNo ratings yet
- Associations of Glucose Control with Insulin Sensitivity and Pancreatic β-Cell Responsiveness in Newly Presenting Type 2 DiabetesDocument6 pagesAssociations of Glucose Control with Insulin Sensitivity and Pancreatic β-Cell Responsiveness in Newly Presenting Type 2 DiabetesJoseph PositivoNo ratings yet
- Defining Insulin Resistance From Hyperinsulinemic-Euglycemic ClampsDocument6 pagesDefining Insulin Resistance From Hyperinsulinemic-Euglycemic ClampsNanda Rizky FathiyaNo ratings yet
- Robust PBPK/PD-Based Model PredictiveDocument13 pagesRobust PBPK/PD-Based Model PredictivenhatvpNo ratings yet
- Serum Irisin With Obesity-Associated Metabolic Syndrome, Type II Diabetes, and Insulin Resistance BackgroundDocument19 pagesSerum Irisin With Obesity-Associated Metabolic Syndrome, Type II Diabetes, and Insulin Resistance BackgroundAmany RadwanNo ratings yet
- Zhou2019 Article InsulinDegludecANovelUltra-LonDocument18 pagesZhou2019 Article InsulinDegludecANovelUltra-LonmehakNo ratings yet
- 10.1515 - CCLM 2023 7047Document101 pages10.1515 - CCLM 2023 7047sfendri17No ratings yet
- Tmp117e TMPDocument6 pagesTmp117e TMPFrontiersNo ratings yet
- Protocolo YaleDocument7 pagesProtocolo Yalegiseladelarosa2006No ratings yet
- Sensors 21 07117Document25 pagesSensors 21 07117fhamdiNo ratings yet
- 2010 TCF7L2 Variant Rs7903146 Affects The Risk of Type 2 Diabetes by Modulating Incretin ActionDocument7 pages2010 TCF7L2 Variant Rs7903146 Affects The Risk of Type 2 Diabetes by Modulating Incretin ActionFiqih NurkholisNo ratings yet
- 2019 Non-invasiveMonitoringOfBloodG ForskinDocument15 pages2019 Non-invasiveMonitoringOfBloodG ForskinhcyNo ratings yet
- Thyroid Disrupting Chemicals Inside Mix Perturb Thymocyte Distinction Throughout Xenopus Laevis TadpolesozdlcDocument1 pageThyroid Disrupting Chemicals Inside Mix Perturb Thymocyte Distinction Throughout Xenopus Laevis Tadpolesozdlccuppantry59No ratings yet
- Changes of Insulin Sensitivity and Secretion After Bariatric Metabolic SurgeryDocument7 pagesChanges of Insulin Sensitivity and Secretion After Bariatric Metabolic SurgeryfrandellionNo ratings yet
- Final Draft - Surya Bagaskara - Identifying Risk Factors of Prediabetes in Prehypertension PopulationDocument10 pagesFinal Draft - Surya Bagaskara - Identifying Risk Factors of Prediabetes in Prehypertension PopulationHerfandi DimasNo ratings yet
- Ajtr0014 4757Document20 pagesAjtr0014 4757Eyal RobinsonNo ratings yet
- 10 1213@ane 0000000000002814Document8 pages10 1213@ane 0000000000002814jaquelinemiyakeNo ratings yet
- Non-Invasive Blood Glucose Level Measurement From LASER Reflected Spectral Patterns ImagesDocument5 pagesNon-Invasive Blood Glucose Level Measurement From LASER Reflected Spectral Patterns ImagesIOSRJEN : hard copy, certificates, Call for Papers 2013, publishing of journalNo ratings yet
- 62 Ej14-0602 PDFDocument11 pages62 Ej14-0602 PDFEsteban Martin Chiotti KaneshimaNo ratings yet
- Effect of Increasing Duration of Diabetes Mellitus Type 2 On Glycated Hemoglobin and Insulin SensitivityDocument5 pagesEffect of Increasing Duration of Diabetes Mellitus Type 2 On Glycated Hemoglobin and Insulin SensitivityAlifLaelaNo ratings yet
- Comparison of The Accuracy of The Hemocue Glucose Analyzer With The Yellow Springs Instrument Glucose Oxidase Analyzer, Particularly in HypoglycemiaDocument7 pagesComparison of The Accuracy of The Hemocue Glucose Analyzer With The Yellow Springs Instrument Glucose Oxidase Analyzer, Particularly in HypoglycemiaSeven NightNo ratings yet
- Metabolism: Effect of Lifestyle Intervention in Patients With Type 2 Diabetes: A Meta-AnalysisDocument10 pagesMetabolism: Effect of Lifestyle Intervention in Patients With Type 2 Diabetes: A Meta-AnalysisAoy RangsimaNo ratings yet
- Synthesis PaperDocument10 pagesSynthesis Paperapi-302383914No ratings yet
- The Cutoff Values of Indirect Indices For Measuring Insulin Resistance For Metabolic Syndrome in Korean Children and AdolescentsDocument6 pagesThe Cutoff Values of Indirect Indices For Measuring Insulin Resistance For Metabolic Syndrome in Korean Children and AdolescentsSathyanarayana SrikantaNo ratings yet
- 10 1152@ajpendo 00512 2019Document52 pages10 1152@ajpendo 00512 2019Aldo González SalinasNo ratings yet
- Original ArticleDocument10 pagesOriginal ArticlegistaluvikaNo ratings yet
- Home Use of An Artificial Beta Cell in Type 1 Diabetes: Original ArticleDocument12 pagesHome Use of An Artificial Beta Cell in Type 1 Diabetes: Original ArticleCarmen UngureanuNo ratings yet
- Generating Molecular Database Using Biocomputing ApproachDocument10 pagesGenerating Molecular Database Using Biocomputing ApproachResearch Cell: An International Journal of Engineering SciencesNo ratings yet
- Wang Et Al., (2018) Relative Validity of An Indirect CalorimetryDocument6 pagesWang Et Al., (2018) Relative Validity of An Indirect CalorimetryAna Flávia SordiNo ratings yet
- Comparison Between PID-Fuzzy and Numerical Methods Based On Linear Algebra Controllers For Glucose Control in Type 1 Diabetes TreatmentDocument7 pagesComparison Between PID-Fuzzy and Numerical Methods Based On Linear Algebra Controllers For Glucose Control in Type 1 Diabetes Treatmentkaveendra.rcitNo ratings yet
- Generating A Reference Interval For Fasting Serum Insulin in Healthy Nondiabetic Adult Chinese MenDocument5 pagesGenerating A Reference Interval For Fasting Serum Insulin in Healthy Nondiabetic Adult Chinese MenTanveerNo ratings yet
- A Mathematical Model of Glucose - Insulin Regulation Under The Influence of Externally Ingested Glucose (G-I-E Model)Document5 pagesA Mathematical Model of Glucose - Insulin Regulation Under The Influence of Externally Ingested Glucose (G-I-E Model)inventionjournalsNo ratings yet
- Mazidi 2017Document15 pagesMazidi 2017Anh Tho PHAMNo ratings yet
- Clinical Course of Steroid Sensitive Nephrotic Syndrome in Children: Outcome and OutlookDocument11 pagesClinical Course of Steroid Sensitive Nephrotic Syndrome in Children: Outcome and OutlookIkhbar FalahNo ratings yet
- Continuous Glucose MonitoringFrom EverandContinuous Glucose MonitoringWeiping JiaNo ratings yet
- 2017 T1DF Glucagon Action - Complaint (Case 2:17-cv-00802-RAJ Filed On May, 24 2017 by Keller Rohrback)Document293 pages2017 T1DF Glucagon Action - Complaint (Case 2:17-cv-00802-RAJ Filed On May, 24 2017 by Keller Rohrback)The Type 1 Diabetes Defense FoundationNo ratings yet
- Diabetes Prevention and Control Program Chapter 5Document24 pagesDiabetes Prevention and Control Program Chapter 5Wendy EscalanteNo ratings yet
- Ac Test 2 Question PaperDocument23 pagesAc Test 2 Question PaperSherylen AlmalbisNo ratings yet
- Chromium For Type 2 DiabetesDocument6 pagesChromium For Type 2 DiabetesSathishKumarNo ratings yet
- Himalayan Salt BenefitsDocument5 pagesHimalayan Salt BenefitsRani John Tayag100% (2)
- AppleDocument13 pagesApplecksharma68No ratings yet
- Coy Diet Patient BrochureDocument32 pagesCoy Diet Patient BrochureRebeca Daniela CherloabaNo ratings yet
- Gliclazide Modified Release.10Document8 pagesGliclazide Modified Release.10Sowbhagya LaxmiNo ratings yet
- Pathophysiology of DiabetesDocument88 pagesPathophysiology of DiabetesCahya SetiyaNo ratings yet
- Diabetes Mellitus - ICD-10 CodingDocument14 pagesDiabetes Mellitus - ICD-10 CodingTim T VargheseNo ratings yet
- FNCP 3Document11 pagesFNCP 3Lucelle Arellano0% (1)
- Diabetes Awareness PamphletDocument3 pagesDiabetes Awareness Pamphletapi-341618058No ratings yet
- Short Guide Clinical NutritionDocument30 pagesShort Guide Clinical Nutritiontopiq100% (1)
- Nutrition Repaired)Document10 pagesNutrition Repaired)Brylle MirandaNo ratings yet
- Fundamentals of Nursing 1Document6 pagesFundamentals of Nursing 1Ruel M. BersabeNo ratings yet
- Paleo Cures: Tota L Hea LTH Acceler AtorDocument9 pagesPaleo Cures: Tota L Hea LTH Acceler AtorLavanya Priya SathyanNo ratings yet
- Natural Diabetes Treatment InformationDocument5 pagesNatural Diabetes Treatment Informationxango2001No ratings yet
- J Diabetes Sci Technol 2009 Ginsberg 903 13Document11 pagesJ Diabetes Sci Technol 2009 Ginsberg 903 13Ninosk Mendoza SolisNo ratings yet
- Pex 04 02Document4 pagesPex 04 02Natalie Cristine Innecken100% (2)
- Type2dm PDFDocument3 pagesType2dm PDFapi-303065684No ratings yet
- Glucometro ProdigyDocument96 pagesGlucometro Prodigyrponce161276No ratings yet
- OSCE S BibleDocument66 pagesOSCE S BibleApoorva SaxenaNo ratings yet
- Graphing ActivitiesDocument12 pagesGraphing Activitiesapi-292104717No ratings yet
- Acute. KetoacidosisdocxDocument12 pagesAcute. KetoacidosisdocxShara SampangNo ratings yet
- Effects of Human Growth Hormone: Process Diagrams Step-by-StepDocument11 pagesEffects of Human Growth Hormone: Process Diagrams Step-by-Steprambabs369100% (1)
- What Is Diabetes? What Causes Diabetes?: Fast Facts On DiabetesDocument6 pagesWhat Is Diabetes? What Causes Diabetes?: Fast Facts On DiabetesAlfred Melvin SolivaNo ratings yet
- Physical Therapy For Cardiopumonary DisordersDocument101 pagesPhysical Therapy For Cardiopumonary Disorderssunita_h100% (5)
- Diabetes and Virgin Coconut Oil DR Bruce FifeDocument3 pagesDiabetes and Virgin Coconut Oil DR Bruce Fifepetri_jv100% (1)
- 5 Steps To Control High Blood SugarDocument56 pages5 Steps To Control High Blood Sugarjha.sofcon5941100% (1)
























































































Download as pdf or txt
You might also like
- SJH Newcastle Diet Booklet 2012 Revised 2015Document6 pagesSJH Newcastle Diet Booklet 2012 Revised 2015TexasPedi100% (1)
- tmp22D3 TMPDocument9 pagestmp22D3 TMPFrontiersNo ratings yet
- RM ProtocolDocument4 pagesRM Protocolcifha diasNo ratings yet
- Thorough Testing of Predictive Functional Control On A Well Validated in Silico Model of Type I Diabetic PatientsDocument12 pagesThorough Testing of Predictive Functional Control On A Well Validated in Silico Model of Type I Diabetic PatientsgercampeNo ratings yet
- Manuscript 24092017 UcldDocument46 pagesManuscript 24092017 UcldPrima Hari NastitiNo ratings yet
- Divya 13Document20 pagesDivya 13Divya NadarNo ratings yet
- Song 2014Document11 pagesSong 2014Ayssa WitjaksonoNo ratings yet
- Journal Pone 0248280 PDFDocument17 pagesJournal Pone 0248280 PDFHafis KafitNo ratings yet
- Measurement of Glucose Homeostasis in VivoDocument10 pagesMeasurement of Glucose Homeostasis in Vivo19400668No ratings yet
- Clinical Study: Obesity and Cytokines in Childhood-Onset Systemic Lupus ErythematosusDocument6 pagesClinical Study: Obesity and Cytokines in Childhood-Onset Systemic Lupus ErythematosusIndra KusumayadiNo ratings yet
- Article AmalDocument16 pagesArticle Amalsaliha mharchiNo ratings yet
- InositolDocument14 pagesInositolEdna De Azevedo GabrielNo ratings yet
- Estimating Insulin Sensitivity and Beta-Cell Function From The Oral Glucose Tolerance Test (Ha Et Al., 2024)Document47 pagesEstimating Insulin Sensitivity and Beta-Cell Function From The Oral Glucose Tolerance Test (Ha Et Al., 2024)Marcelo TrNo ratings yet
- Association of Coagulation Profile With Microvascular Complications and Glycemic Control in Type 2 Diabetes Mellitus - A Study ADocument1 pageAssociation of Coagulation Profile With Microvascular Complications and Glycemic Control in Type 2 Diabetes Mellitus - A Study Aseneh17198No ratings yet
- Continuous Glucose Monitoring Devices For Patients With Diabeles Mellitus On InsulinDocument25 pagesContinuous Glucose Monitoring Devices For Patients With Diabeles Mellitus On InsulinCalifornia Technology Assessment ForumNo ratings yet
- 11 Effect Probiotics On Glucose MetabolismDocument7 pages11 Effect Probiotics On Glucose Metabolismbangd1f4nNo ratings yet
- Diabetes Research and Clinical Practice Volume Issue 2018Document37 pagesDiabetes Research and Clinical Practice Volume Issue 2018yayan_adiutama98No ratings yet
- First-Trimester Prediction of Gestational Diabetes Mellitus: Examining The Potential of Combining Maternal Characteristics and Laboratory MeasuresDocument6 pagesFirst-Trimester Prediction of Gestational Diabetes Mellitus: Examining The Potential of Combining Maternal Characteristics and Laboratory MeasuresLucky PratamaNo ratings yet
- Glycemic 2010 PDFDocument9 pagesGlycemic 2010 PDFSiva SankarNo ratings yet
- Glutamina IVDocument11 pagesGlutamina IVBLADIMIR ALEJANDRO GIL VALENCIANo ratings yet
- Human Insulin Vs Insulin AnalogDocument7 pagesHuman Insulin Vs Insulin AnalogWita Ferani Kartika100% (1)
- Tight Glycemic Control in Critically Ill PediatricDocument11 pagesTight Glycemic Control in Critically Ill Pediatricbella friscaamaliaNo ratings yet
- Gene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GeneDocument7 pagesGene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GenesaifudinNo ratings yet
- Associations Between The MetAssociations Between The Metabolic Syndrome Abolic Syndrome andDocument9 pagesAssociations Between The MetAssociations Between The Metabolic Syndrome Abolic Syndrome andEsteban Avila IraguenNo ratings yet
- Time-Delay Model-Based Control of The Glucose-Insulin System, by Means of A State ObserverDocument16 pagesTime-Delay Model-Based Control of The Glucose-Insulin System, by Means of A State ObserverChanachai PuckNo ratings yet
- Palumbo 13Document13 pagesPalumbo 13Margarete Ap SantosNo ratings yet
- Associations of Glucose Control with Insulin Sensitivity and Pancreatic β-Cell Responsiveness in Newly Presenting Type 2 DiabetesDocument6 pagesAssociations of Glucose Control with Insulin Sensitivity and Pancreatic β-Cell Responsiveness in Newly Presenting Type 2 DiabetesJoseph PositivoNo ratings yet
- Defining Insulin Resistance From Hyperinsulinemic-Euglycemic ClampsDocument6 pagesDefining Insulin Resistance From Hyperinsulinemic-Euglycemic ClampsNanda Rizky FathiyaNo ratings yet
- Robust PBPK/PD-Based Model PredictiveDocument13 pagesRobust PBPK/PD-Based Model PredictivenhatvpNo ratings yet
- Serum Irisin With Obesity-Associated Metabolic Syndrome, Type II Diabetes, and Insulin Resistance BackgroundDocument19 pagesSerum Irisin With Obesity-Associated Metabolic Syndrome, Type II Diabetes, and Insulin Resistance BackgroundAmany RadwanNo ratings yet
- Zhou2019 Article InsulinDegludecANovelUltra-LonDocument18 pagesZhou2019 Article InsulinDegludecANovelUltra-LonmehakNo ratings yet
- 10.1515 - CCLM 2023 7047Document101 pages10.1515 - CCLM 2023 7047sfendri17No ratings yet
- Tmp117e TMPDocument6 pagesTmp117e TMPFrontiersNo ratings yet
- Protocolo YaleDocument7 pagesProtocolo Yalegiseladelarosa2006No ratings yet
- Sensors 21 07117Document25 pagesSensors 21 07117fhamdiNo ratings yet
- 2010 TCF7L2 Variant Rs7903146 Affects The Risk of Type 2 Diabetes by Modulating Incretin ActionDocument7 pages2010 TCF7L2 Variant Rs7903146 Affects The Risk of Type 2 Diabetes by Modulating Incretin ActionFiqih NurkholisNo ratings yet
- 2019 Non-invasiveMonitoringOfBloodG ForskinDocument15 pages2019 Non-invasiveMonitoringOfBloodG ForskinhcyNo ratings yet
- Thyroid Disrupting Chemicals Inside Mix Perturb Thymocyte Distinction Throughout Xenopus Laevis TadpolesozdlcDocument1 pageThyroid Disrupting Chemicals Inside Mix Perturb Thymocyte Distinction Throughout Xenopus Laevis Tadpolesozdlccuppantry59No ratings yet
- Changes of Insulin Sensitivity and Secretion After Bariatric Metabolic SurgeryDocument7 pagesChanges of Insulin Sensitivity and Secretion After Bariatric Metabolic SurgeryfrandellionNo ratings yet
- Final Draft - Surya Bagaskara - Identifying Risk Factors of Prediabetes in Prehypertension PopulationDocument10 pagesFinal Draft - Surya Bagaskara - Identifying Risk Factors of Prediabetes in Prehypertension PopulationHerfandi DimasNo ratings yet
- Ajtr0014 4757Document20 pagesAjtr0014 4757Eyal RobinsonNo ratings yet
- 10 1213@ane 0000000000002814Document8 pages10 1213@ane 0000000000002814jaquelinemiyakeNo ratings yet
- Non-Invasive Blood Glucose Level Measurement From LASER Reflected Spectral Patterns ImagesDocument5 pagesNon-Invasive Blood Glucose Level Measurement From LASER Reflected Spectral Patterns ImagesIOSRJEN : hard copy, certificates, Call for Papers 2013, publishing of journalNo ratings yet
- 62 Ej14-0602 PDFDocument11 pages62 Ej14-0602 PDFEsteban Martin Chiotti KaneshimaNo ratings yet
- Effect of Increasing Duration of Diabetes Mellitus Type 2 On Glycated Hemoglobin and Insulin SensitivityDocument5 pagesEffect of Increasing Duration of Diabetes Mellitus Type 2 On Glycated Hemoglobin and Insulin SensitivityAlifLaelaNo ratings yet
- Comparison of The Accuracy of The Hemocue Glucose Analyzer With The Yellow Springs Instrument Glucose Oxidase Analyzer, Particularly in HypoglycemiaDocument7 pagesComparison of The Accuracy of The Hemocue Glucose Analyzer With The Yellow Springs Instrument Glucose Oxidase Analyzer, Particularly in HypoglycemiaSeven NightNo ratings yet
- Metabolism: Effect of Lifestyle Intervention in Patients With Type 2 Diabetes: A Meta-AnalysisDocument10 pagesMetabolism: Effect of Lifestyle Intervention in Patients With Type 2 Diabetes: A Meta-AnalysisAoy RangsimaNo ratings yet
- Synthesis PaperDocument10 pagesSynthesis Paperapi-302383914No ratings yet
- The Cutoff Values of Indirect Indices For Measuring Insulin Resistance For Metabolic Syndrome in Korean Children and AdolescentsDocument6 pagesThe Cutoff Values of Indirect Indices For Measuring Insulin Resistance For Metabolic Syndrome in Korean Children and AdolescentsSathyanarayana SrikantaNo ratings yet
- 10 1152@ajpendo 00512 2019Document52 pages10 1152@ajpendo 00512 2019Aldo González SalinasNo ratings yet
- Original ArticleDocument10 pagesOriginal ArticlegistaluvikaNo ratings yet
- Home Use of An Artificial Beta Cell in Type 1 Diabetes: Original ArticleDocument12 pagesHome Use of An Artificial Beta Cell in Type 1 Diabetes: Original ArticleCarmen UngureanuNo ratings yet
- Generating Molecular Database Using Biocomputing ApproachDocument10 pagesGenerating Molecular Database Using Biocomputing ApproachResearch Cell: An International Journal of Engineering SciencesNo ratings yet
- Wang Et Al., (2018) Relative Validity of An Indirect CalorimetryDocument6 pagesWang Et Al., (2018) Relative Validity of An Indirect CalorimetryAna Flávia SordiNo ratings yet
- Comparison Between PID-Fuzzy and Numerical Methods Based On Linear Algebra Controllers For Glucose Control in Type 1 Diabetes TreatmentDocument7 pagesComparison Between PID-Fuzzy and Numerical Methods Based On Linear Algebra Controllers For Glucose Control in Type 1 Diabetes Treatmentkaveendra.rcitNo ratings yet
- Generating A Reference Interval For Fasting Serum Insulin in Healthy Nondiabetic Adult Chinese MenDocument5 pagesGenerating A Reference Interval For Fasting Serum Insulin in Healthy Nondiabetic Adult Chinese MenTanveerNo ratings yet
- A Mathematical Model of Glucose - Insulin Regulation Under The Influence of Externally Ingested Glucose (G-I-E Model)Document5 pagesA Mathematical Model of Glucose - Insulin Regulation Under The Influence of Externally Ingested Glucose (G-I-E Model)inventionjournalsNo ratings yet
- Mazidi 2017Document15 pagesMazidi 2017Anh Tho PHAMNo ratings yet
- Clinical Course of Steroid Sensitive Nephrotic Syndrome in Children: Outcome and OutlookDocument11 pagesClinical Course of Steroid Sensitive Nephrotic Syndrome in Children: Outcome and OutlookIkhbar FalahNo ratings yet
- Continuous Glucose MonitoringFrom EverandContinuous Glucose MonitoringWeiping JiaNo ratings yet
- 2017 T1DF Glucagon Action - Complaint (Case 2:17-cv-00802-RAJ Filed On May, 24 2017 by Keller Rohrback)Document293 pages2017 T1DF Glucagon Action - Complaint (Case 2:17-cv-00802-RAJ Filed On May, 24 2017 by Keller Rohrback)The Type 1 Diabetes Defense FoundationNo ratings yet
- Diabetes Prevention and Control Program Chapter 5Document24 pagesDiabetes Prevention and Control Program Chapter 5Wendy EscalanteNo ratings yet
- Ac Test 2 Question PaperDocument23 pagesAc Test 2 Question PaperSherylen AlmalbisNo ratings yet
- Chromium For Type 2 DiabetesDocument6 pagesChromium For Type 2 DiabetesSathishKumarNo ratings yet
- Himalayan Salt BenefitsDocument5 pagesHimalayan Salt BenefitsRani John Tayag100% (2)
- AppleDocument13 pagesApplecksharma68No ratings yet
- Coy Diet Patient BrochureDocument32 pagesCoy Diet Patient BrochureRebeca Daniela CherloabaNo ratings yet
- Gliclazide Modified Release.10Document8 pagesGliclazide Modified Release.10Sowbhagya LaxmiNo ratings yet
- Pathophysiology of DiabetesDocument88 pagesPathophysiology of DiabetesCahya SetiyaNo ratings yet
- Diabetes Mellitus - ICD-10 CodingDocument14 pagesDiabetes Mellitus - ICD-10 CodingTim T VargheseNo ratings yet
- FNCP 3Document11 pagesFNCP 3Lucelle Arellano0% (1)
- Diabetes Awareness PamphletDocument3 pagesDiabetes Awareness Pamphletapi-341618058No ratings yet
- Short Guide Clinical NutritionDocument30 pagesShort Guide Clinical Nutritiontopiq100% (1)
- Nutrition Repaired)Document10 pagesNutrition Repaired)Brylle MirandaNo ratings yet
- Fundamentals of Nursing 1Document6 pagesFundamentals of Nursing 1Ruel M. BersabeNo ratings yet
- Paleo Cures: Tota L Hea LTH Acceler AtorDocument9 pagesPaleo Cures: Tota L Hea LTH Acceler AtorLavanya Priya SathyanNo ratings yet
- Natural Diabetes Treatment InformationDocument5 pagesNatural Diabetes Treatment Informationxango2001No ratings yet
- J Diabetes Sci Technol 2009 Ginsberg 903 13Document11 pagesJ Diabetes Sci Technol 2009 Ginsberg 903 13Ninosk Mendoza SolisNo ratings yet
- Pex 04 02Document4 pagesPex 04 02Natalie Cristine Innecken100% (2)
- Type2dm PDFDocument3 pagesType2dm PDFapi-303065684No ratings yet
- Glucometro ProdigyDocument96 pagesGlucometro Prodigyrponce161276No ratings yet
- OSCE S BibleDocument66 pagesOSCE S BibleApoorva SaxenaNo ratings yet
- Graphing ActivitiesDocument12 pagesGraphing Activitiesapi-292104717No ratings yet
- Acute. KetoacidosisdocxDocument12 pagesAcute. KetoacidosisdocxShara SampangNo ratings yet
- Effects of Human Growth Hormone: Process Diagrams Step-by-StepDocument11 pagesEffects of Human Growth Hormone: Process Diagrams Step-by-Steprambabs369100% (1)
- What Is Diabetes? What Causes Diabetes?: Fast Facts On DiabetesDocument6 pagesWhat Is Diabetes? What Causes Diabetes?: Fast Facts On DiabetesAlfred Melvin SolivaNo ratings yet
- Physical Therapy For Cardiopumonary DisordersDocument101 pagesPhysical Therapy For Cardiopumonary Disorderssunita_h100% (5)
- Diabetes and Virgin Coconut Oil DR Bruce FifeDocument3 pagesDiabetes and Virgin Coconut Oil DR Bruce Fifepetri_jv100% (1)
- 5 Steps To Control High Blood SugarDocument56 pages5 Steps To Control High Blood Sugarjha.sofcon5941100% (1)